
Ст 133 ч 3 тк рф: Статья 133 [ТК РФ] – последняя редакция
Об оплате труда работника, выполняющего работу дистанционно
В силу положений Трудового кодекса РФ (ст. 15, ч. 1 ст. 56 ТК РФ) трудовые отношения носят возмездный характер.
Получение своевременной и в полном объеме заработной платы является одним из основных прав работника, а своевременная и в полном размере ее выплата – главной обязанностью работодателя (абз. 5 ч. 1 ст. 21, абз. 7 ч. 2 ст. 22 ТК РФ).
При этом в случае выполнения работником работы никакие внешние факторы – чрезвычайные обстоятельства, бедствия или угрозы бедствия (пожары, наводнения, голод, землетрясения, эпидемии или эпизоотии) и иные случаи, ставящие под угрозу жизнь или нормальные жизненные условия всего населения или его части, не должны препятствовать реализации этого права и обязанности.
Если работодатель в целях противодействия угрозе распространения коронавирусной инфекции принял решение перевести всех своих работников или часть из них на дистанционную работу, то он должен учитывать, что на дистанционных работников распространяется действие трудового законодательства и иных актов, содержащих нормы трудового права.
 3 ст. 133 ТК РФ).
3 ст. 133 ТК РФ).В соответствии с требованиями Трудового кодекса (ст. 15, ч. 1 ст. 56 ТК РФ) трудовые отношения носят возмездный характер
Получение своевременной и в полном объеме заработной платы является одним из ключевых прав работника, а своевременная и в полном размере ее выплата – главной обязанностью работодателя (абз. 5 ч. 1 ст. 21, абз. 7 ч. 2 ст. 22 ТК РФ).
В случае выполнения работником работы никакие внешние факторы – чрезвычайные обстоятельства, бедствия или угрозы бедствия (пожары, наводнения, голод, землетрясения или эпидемии) и иные случаи, ставящие под угрозу жизнь или нормальные жизненные условия всего населения или его части, не должны препятствовать реализации этого права и обязанности.
Если работодатель принял решение перевести всех своих работников или часть из них на дистанционную работу, то он должен учитывать, что на дистанционных работников распространяется действие трудового законодательства и иных актов, содержащих нормы трудового права.
Поскольку перевод на дистанционный режим носит вынужденный и временный характер, а специфика оплаты труда в такой ситуации ТК РФ прямо не определена, то при сохранении объема трудовых обязанностей не должен измениться и размер оплаты труда. То есть, если фактически корректируется только место работы, а все остальные условия трудового договора продолжают действовать в прежнем виде, зарплата должна выплачиваться в размере, установленном ранее – в период работы в офисе (с учетом всех ее составляющих – оклада, доплат, надбавок, премий, иных компенсационных и стимулирующих выплат).
Однако по договоренности работника с работодателем условия оплаты труда могут быть скорректированы в дополнительном соглашении к трудовому договору, которым урегулированы условия дистанционной работы.
Соответственно месячная заработная плата работника, полностью отработавшего за соответствующий период норму рабочего времени и выполнившего нормы труда (трудовые обязанности), не может быть ниже минимального размера оплаты труда (ч. 3 ст. 133 ТК РФ).
3 ст. 133 ТК РФ).
Тимирязевский межрайонный прокурор А.Г.Захаров
Администрация Шелеховского района. О начислении выплат сверх МРОТ
3 октября 2019 г.
О начислении выплат сверх МРОТ
Конституция Российской Федерации провозглашает Россию правовым социальным государством, политика которого направлена на создание условий, обеспечивающих достойную жизнь и свободное развитие человека, закрепляет, что в Российской Федерации охраняются труд и здоровье людей, каждый имеет право на вознаграждение за труд без какой бы то ни было дискриминации и не ниже установленного Федеральным законом минимального размера оплаты труда (МРОТ).

Согласно ст. 129 Трудового кодекса, заработной платой (оплатой труда работника) признаются вознаграждение за труд в зависимости от квалификации работника, сложности, количества, качества и условий выполняемой работы, а также компенсационные выплаты и стимулирующие выплаты.
Частью 1 ст. 133 ТК РФ предусмотрено, что МРОТ устанавливается одновременно на всей территории Российской Федерации федеральным законом и не может быть ниже величины прожиточного минимума трудоспособного населения. Месячная заработная плата работника, полностью отработавшего за этот период норму рабочего времени и выполнившего нормы труда (трудовые обязанности), не может быть ниже МРОТ (ч. 3 ст. 133 ТК РФ).
В силу ст. 133.1 ТК РФ в субъекте РФ региональным соглашением может устанавливаться размер минимальной заработной платы для работников, работающих на его территории, за исключением работников организаций, финансируемых из федерального бюджета. Размер минимальной заработной платы в субъекте Российской Федерации не может быть ниже МРОТ, установленного федеральным законом.
 Месячная заработная плата работника, работающего на территории соответствующего субъекта РФ и состоящего в трудовых отношениях с работодателем, в отношении которого действует региональное соглашение о минимальной заработной плате, не может быть ниже размера минимальной заработной платы в этом субъекте Российской Федерации при условии, что таким работником полностью отработана за этот период норма рабочего времени и выполнены нормы труда.
Месячная заработная плата работника, работающего на территории соответствующего субъекта РФ и состоящего в трудовых отношениях с работодателем, в отношении которого действует региональное соглашение о минимальной заработной плате, не может быть ниже размера минимальной заработной платы в этом субъекте Российской Федерации при условии, что таким работником полностью отработана за этот период норма рабочего времени и выполнены нормы труда.М.В. СИБГАТУЛИНА,
помощник прокурора города Шелехова
Статья 133 ТК РФ с комментариями.
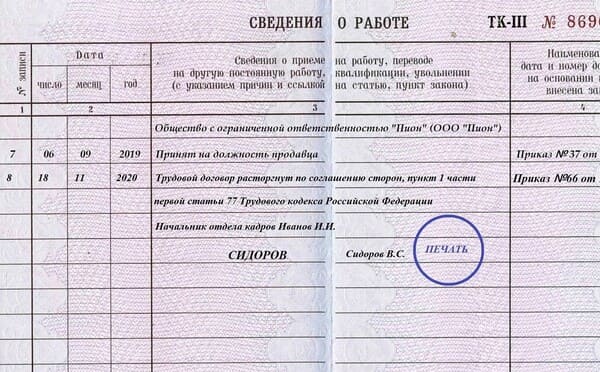 Установление минимального размера оплаты труда (в ред. Федерального закона от 20.04.2007 N 54-ФЗ)
Установление минимального размера оплаты труда (в ред. Федерального закона от 20.04.2007 N 54-ФЗ)(в ред. Федерального закона от 22.08.2004 N 122-ФЗ)
КонсультантПлюс: примечание.
Порядок и сроки поэтапного повышения МРОТ устанавливаются федеральным законом (ст. 421 данного документа).
КонсультантПлюс: примечание.
О выявлении конституционно-правового смысла ч. 1 ст. 133 см. Постановления КС РФ от 11.04.2019 N 17-П, от 16.12.2019 N 40-П.
Минимальный размер оплаты труда устанавливается одновременно на всей территории Российской Федерации федеральным законом и не может быть ниже величины прожиточного минимума трудоспособного населения.
(в ред. Федеральных законов от 30.06.2006 N 90-ФЗ, от 20.04.2007 N 54-ФЗ)
Минимальный размер оплаты труда, установленный федеральным законом, обеспечивается:
организациями, финансируемыми из федерального бюджета, – за счет средств федерального бюджета, внебюджетных средств, а также средств, полученных от предпринимательской и иной приносящей доход деятельности;
(в ред. Федерального закона от 20.04.2007 N 54-ФЗ)
Федерального закона от 20.04.2007 N 54-ФЗ)
организациями, финансируемыми из бюджетов субъектов Российской Федерации, – за счет средств бюджетов субъектов Российской Федерации, внебюджетных средств, а также средств, полученных от предпринимательской и иной приносящей доход деятельности;
(в ред. Федерального закона от 20.04.2007 N 54-ФЗ)
организациями, финансируемыми из местных бюджетов, – за счет средств местных бюджетов, внебюджетных средств, а также средств, полученных от предпринимательской и иной приносящей доход деятельности;
(в ред. Федерального закона от 20.04.2007 N 54-ФЗ)
другими работодателями – за счет собственных средств.
(часть вторая в ред. Федерального закона от 30.06.2006 N 90-ФЗ)
КонсультантПлюс: примечание.
О выявлении конституционно-правового смысла ч. 3 ст. 133 см. Постановления КС РФ от 11.04.2019 N 17-П, от 16.12.2019 N 40-П.
Месячная заработная плата работника, полностью отработавшего за этот период норму рабочего времени и выполнившего нормы труда (трудовые обязанности), не может быть ниже минимального размера оплаты труда.
(в ред. Федеральных законов от 30.06.2006 N 90-ФЗ, от 20.04.2007 N 54-ФЗ)
Часть четвертая утратила силу с 1 сентября 2007 года. – Федеральный закон от 20.04.2007 N 54-ФЗ.
Чем режим нерабочих дней отличается от выходных и как он оплачивается? — Справки
Выплата зарплаты в праздничные и выходные дни устанавливается коллективным или трудовым договором (ст. 149 ТК РФ). Размер таких доплат может быть любым, но не ниже минимального уровня, который указан в статье 153 ТК. Если работник выходит на работу в такой день по инициативе работодателя, тот этот день должен быть оплачен как минимум в двойном размере (ст. 153 ТК РФ). Если сотрудник отработает в праздничный день несколько часов, ему заплатят в двойном размере только за эти часы, а не за полный день (ч. 3 ст. 153 ТК).
Читайте также
Никаких «нерабочих дней с сохранением заработной платы» в российском трудовом законодательстве не прописано. Каждый раз вопрос о формате оплаты таких дней указывается в конкретном указе или постановлении, говорит Ольга Петрова.
«За праздничные дни мы все получаем зарплату, а выходные дни у всех разные. У кого-то выходной в субботу и в воскресенье, а у кого-то, кто работает по другому режиму, это могут быть другие дни — и среды, и четверги и понедельники. Режим нерабочих дней означает, что это нерабочие дни, за которые по обычному графику мы не получаем деньги, но если принимается решение и в принятом документе указано, что за такие дни предусмотрена оплата вне зависимости от того, что они нерабочие, то тогда это просто дополнительные условия к указу. В Трудовом кодексе такого режима нет. Но в указе может быть прописано, что это будет нерабочий день, и за него деньги вы все равно получите. Если оплата за такой день не прописана и вас привлекают к труду в связи с невозможностью остановить производство и т. д., то работа в такой день оплачивается. Все условия прописываются в документе», —говорит Петрова.
Таким образом, это может быть просто нерабочий день или нерабочий день с сохранением заработной платы.
Ольга Петрова
эксперт по трудовому праву

О том, что предприятия не должны снижать зарплату в связи с нерабочими днями и обязаны выплатить ее сотрудникам в полном объеме, ранее заявляли в правительстве. В Минтруде поясняли, что оплату нерабочих дней рекомендовано производить в обычном, а не повышенном размере, если работодатель не решит иначе.
Нерабочие дни согласно прошлым указам президента не относили к выходным или нерабочим праздничным дням, поэтому они не уменьшали норму рабочего времени. (письмо Минтруда от 26.03.2020 № 14−4/10/П-2696).
постановка на учет, отчетность, проводки
Также автор рассказывает какую отчетность необходимо сдавать предприятиям имеющие обособленное подразделение, бухгалтерские проводки необходимые для учета хозяйственных деятельности.
Про обособленные подразделения слышали наверно все, но все же у некоторых руководителей и бухгалтеров возникают вопросы по их открытию, как вести бухгалтерский и налоговый учет, как отчитываться перед ИФНС при их наличии.
Итак, начнем с определения, как бы это скучно не казалось. Обособленное подразделение (далее по тексту ОП) согласно п. 2 ст. 11 НК РФ это любое территориально обособленное от организации подразделение, по месту нахождения которого оборудованы стационарные рабочие места, акцентирую ваше внимание стационарные (например вы сняли помещение для склада в соседнем районе или открыли магазинчик на соседней улице или в соседнем здании в результате таких манипуляций образуется ОП, то есть вы организовали стационарное рабочее место) в этом случае вы (как головная организация) только сообщаете( посредством уведомления) в налоговую инспекцию об открытии ОП, в результате этих действий ИФНС ставит на налоговый учет ОП, запись в ЕГРЮЛ при этом не производится п. 2 ст. 23, п. 4 ст. 83 НК РФ, не путайте с филиалом. Напомню, что определение рабочего места дано в ст. 209 ТК РФ, то есть это место, где работник должен находиться или куда ему необходимо прибыть в связи с его работой и которое прямо или косвенно находится под контролем работодателя. Стационарное место должно быть оборудовано более чем на 1 месяц.
Стационарное место должно быть оборудовано более чем на 1 месяц.
Очень много непонимания возникает у строительных организаций о необходимости образования ОП. Здесь надо исходить из следующих показателей. Если генподрядчик выполняет работы собственными силами или направляет своих работников в командировки на объект строительства (подтверждается табелями учета рабочего времени, командировочными удостоверениями, служебными заданиями) для отслеживания работ субподрядчиков, в результате он создает рабочие места более 1 месяца в этом случае необходимо известить налоговую инспекцию об образовании ОП.
В Постановлении Девятнадцатого арбитражного апелляционного суда от 27.02.2014 по делу N А64-5102/2013, при выполнении строительных работ без образования ОП, суд руководствовался определениями стационарного и нестационарного рабочего места (Постановлением Госстроя России от 31.03.2000 N 26 принят и введен в действие СП 12-133-2000 “Безопасность труда в строительстве. Положение о порядке аттестации рабочих мест по условиям труда в строительстве и жилищно-коммунальном хозяйстве”). Судом был сделан вывод, что для выполнения работ согласно договорам подряда и субподряда на выполнение строительных работ, оборудованы нестационарные рабочие места. Что, в результате, не приводит к образованию обособленного подразделения согласно п. 2 ст. 11 НК РФ, в котором говорится о стационарном рабочем месте.
Судом был сделан вывод, что для выполнения работ согласно договорам подряда и субподряда на выполнение строительных работ, оборудованы нестационарные рабочие места. Что, в результате, не приводит к образованию обособленного подразделения согласно п. 2 ст. 11 НК РФ, в котором говорится о стационарном рабочем месте.
Что же такое нестационарное место — это рабочее место, месторасположение и техническое оснащение которого имеют нестационарный характер, то есть рабочее место связано с определенным строительным объектом или эксплуатируемым сооружением, а техническое оснащение является мобильным или переносным.
Поэтому при организации ОП вам необходимо все тщательно изучить, например тот факт, находятся ли лица, привлекаемые на строительные работы с генподрядчиком в трудовых отношениях или нет. Помните, что доначисление налогов будет существенным если не сообщить вовремя об образовании ОП в налоговый орган. Заявление о постановке на учет подается по форме № 1-2‑Учет в течение одного месяца со дня создания ОП в налоговый орган по месту его нахождения (п. 4 ст. 83 НК РФ). Также по месту своего нахождения налогоплательщики-организации обязаны письменно сообщать обо всех ОП, созданных на территории РФ, в течение одного месяца со дня создания по форме № С-09-3 «Сообщение о создании (закрытии) на территории РФ обособленного подразделения» (пп. 3 п. 2 ст. 23 НК РФ).
4 ст. 83 НК РФ). Также по месту своего нахождения налогоплательщики-организации обязаны письменно сообщать обо всех ОП, созданных на территории РФ, в течение одного месяца со дня создания по форме № С-09-3 «Сообщение о создании (закрытии) на территории РФ обособленного подразделения» (пп. 3 п. 2 ст. 23 НК РФ).
Идем дальше, при образовании ОП, вы как головная организация, можете выделить его на отдельный баланс, а можете вести его без него. Это связано с тем, что для управления ОП руководителям обычно нужны определённые показали (то есть управленческий учет) которые нужны для принятия, оперативных решений, которые в комплексе влияют на показатели всей организации.
Теперь давайте рассмотрим куда необходимо платить налоги и какие.
- Если работник напрямую (то есть по трудовому договору) принят в ОП, НДФЛ с доходов таких работников нужно платить по месту нахождения ОП. Согласно Письма Минфина от 13.08.2018 N 03-04-06/57140 отчеты 2-НДФЛ и 6-НДФЛ по таким работникам сдаются в ИФНС по месту регистрации ОП.
 Но есть оговорка, если ОП несколько и они зарегистрировано в одном муниципальном образовании (районе) необходимо выбрать ответственное ОП которое будет сдавать отчетность за все ОП зарегистрированные в данном районе.
Но есть оговорка, если ОП несколько и они зарегистрировано в одном муниципальном образовании (районе) необходимо выбрать ответственное ОП которое будет сдавать отчетность за все ОП зарегистрированные в данном районе. - ОП платит страховые взносы и сдает отчетность, при условии, что если оно само начисляет и перечисляет выплаты работникам и у него открыт расчетный счет в банке (п. 11 ст. 431 НК РФ).
- НДС ОП не оплачивает и декларацию не сдает, это делает только головная организация в комплексе по всему предприятию:
- уплата и расчет налога на прибыль производится по месту нахождения организации, а также по месту нахождения каждого из ее обособленных подразделений исходя из доли прибыли, приходящейся на эти ОП. Если налогоплательщик имеет несколько ОП на территории одного муниципального образования (района), то распределение прибыли по каждому из этих подразделений может не производиться. Сумма налога, подлежащая уплате в бюджет, в таком случае определяется исходя из доли прибыли, исчисленной из совокупности показателей ОП, находящихся на территории субъекта РФ. Налоговые декларации необходимо представлять в налоговые органы по месту своего нахождения и месту нахождения каждого обособленного подразделения ОП.
- если головная организация находится на УСН и имеет ОП, то сдача декларации и уплата налога производит только головной организацией.
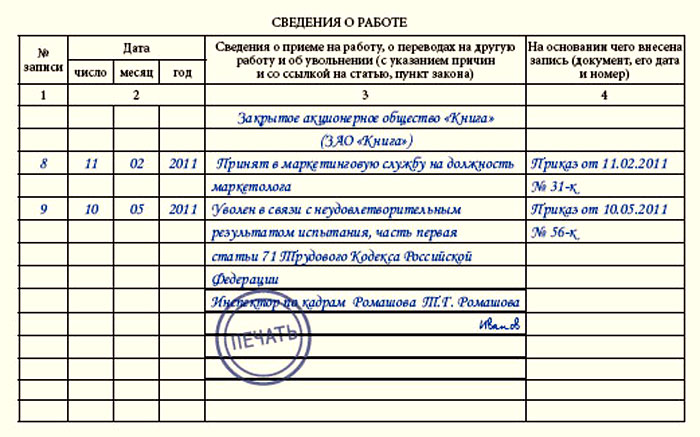 Но есть оговорка, если ОП несколько и они зарегистрировано в одном муниципальном образовании (районе) необходимо выбрать ответственное ОП которое будет сдавать отчетность за все ОП зарегистрированные в данном районе.
Но есть оговорка, если ОП несколько и они зарегистрировано в одном муниципальном образовании (районе) необходимо выбрать ответственное ОП которое будет сдавать отчетность за все ОП зарегистрированные в данном районе.  Налоговые декларации необходимо представлять в налоговые органы по месту своего нахождения и месту нахождения каждого обособленного подразделения ОП.
Налоговые декларации необходимо представлять в налоговые органы по месту своего нахождения и месту нахождения каждого обособленного подразделения ОП.И напоследок разберемся как вести бухгалтерский учет.
Как говорилось ранее можно выделить на отдельном балансе, а можно не выделять. Как принять решение, это зависит что необходимо Вам и Вашему руководителю. Давайте поэтапно.
Учитывая ч. 6 ст. 13 Закона от 06.12.2011 N 402-ФЗ бухгалтерская (финансовая) отчетность организации должна включать показатели деятельности всех ОП, включая выделенные на отдельный баланс. В Письме Минфина от 02.06.2005 N 03-06-01-04/273 дана формулировка что такое отдельный баланс. Итак, под отдельным балансом следует понимать перечень показателей, установленных организацией для своих подразделений, отражающих ее имущественное и финансовое положение на отчетную дату для нужд управления организацией. Отдельный баланс ОП является внутренним документом организации, в налоговую не сдается.
Отдельный баланс ОП является внутренним документом организации, в налоговую не сдается.
Учитывая п. 9 ПБУ 1/2008 «Учетная политика организации», в подразделении, которое выделено на отдельный баланс, применяет единую учетную политику, утвержденную в организации. Порядок отражения в учете взаиморасчетов между ОП и головной организацией, порядок документооборота утверждаются руководителем организации (п. 4 ПБУ 1/2008 «Учетная политика организации»).
Для расчетов с ОП, которые выделены на отдельный баланс, применяется счет 79 «Внутрихозяйственные расчеты». Данный счет используется при передаче ОП какого-либо имущества, при взаимном отпуске материальных ценностей, при оплате труда работников обособленного подразделения и т.д.
Например, если головная организация передает ОП основные средства (ОС), то проводки будут следующие:
- Д 79-1 / К 01 — на первоначальную стоимость объекта;
- Д 02 / К 79-1 – на сумму накопленной амортизации по объекту.
В свою очередь, ОП сделает записи при получении этого ОС:
- Д 01 / К 79-1 — на первоначальную стоимость объекта;
- Д 79-1 / К 02 – на сумму накопленной амортизации по объекту.

Расчеты по счету 79 при составлении единой бухгалтерской отчетности организации не отражаются (сворачиваются).
Эксперт — Обвинцева Екатерина Юрьевна
Президент НП «Ассоциация Профессиональных Бухгалтеров Содружество» Лысенко Д. В.
Как открыть домашний детский сад
Анастасия Макарова
разобралась в детских садах
Профиль автораУ меня трое детей. Моей младшей дочери 3 года, и я задумываюсь о том, чтобы не отдавать ее в обычный детский сад, а организовать свой — домашний.
В статье расскажу, что это такое, чем домашний детский сад отличается от государственного и частного, как организовать все это самому и можно ли получить помощь от государства.
Что такое домашний детский сад
Официально понятия «домашний детский сад» не существует — оно нигде законодательно не закреплено. Но в быту так называют любую форму присмотра за детьми в домашней обстановке. Например, когда одна или несколько мам собирают группу детей и сидят с ними, пока остальные родители на работе. Я сама водила старшего ребенка в такую группу в Санкт-Петербурге 10 лет назад.
Я сама водила старшего ребенка в такую группу в Санкт-Петербурге 10 лет назад.
Часто такие группы в квартирах работают без юридического оформления. Это рискованно и для организаторов, и для родителей: организатору можно получить штраф за нелегальную предпринимательскую деятельность, а еще «воспитатели» в нелегальных садиках не несут никакой ответственности за детей.
ст. 14.1 КоАП РФ
Варианты легального домашнего присмотра за детьми — это официально зарегистрированные частные детские учреждения — группы временного пребывания, развивающие центры, расположенные в жилых помещениях, или семейные дошкольные группы, прикрепленные к государственному учебному заведению.
Семейные дошкольные группы могут организовывать только многодетные родители с детьми-дошкольниками. Называть их можно по-разному — семейными детскими садами как в Москве, группами дошкольного воспитания как в Красноярском крае, домашними детскими садами или иначе.
Частные детские учреждения могут вести образовательную деятельность — в этом случае им нужно иметь лицензию. А могут не заниматься образованием и выполнять другие функции, например присматривать за детьми, кормить их, играть, гулять. В этом случае лицензия не нужна.
А могут не заниматься образованием и выполнять другие функции, например присматривать за детьми, кормить их, играть, гулять. В этом случае лицензия не нужна.
Расскажу обо всех этих вариантах подробнее.
УЧЕБНИК
Как победить выгорание
Курс для тех, кто много работает и устает. Цена открыта — назначаете ее сами
Начать учитьсяРазновидности детских дошкольных заведений
Все дошкольные учреждения в России можно разделить на государственные и частные. Среди государственных выделяются семейные дошкольные группы, а у частных множество разновидностей в зависимости от концепции бизнеса.
Семейная дошкольная группа, или семейный детский сад, — это небольшая группа, в которой воспитатель — это родитель одного или нескольких дошкольников. В такой группе, как правило, должно быть не меньше трех детей — точные требования зависят от региона. Организовать семейную группу могут только многодетные семьи, где есть хотя бы один дошкольник. Дети занимаются у себя дома, но официально прикреплены к муниципальному детскому саду.
Дети занимаются у себя дома, но официально прикреплены к муниципальному детскому саду.
В каждом регионе действуют свои правила по организации таких садов. Например, в Москве открыть семейный детский сад может многодетная семья по месту своего проживания. В семье при этом должно быть от трех детей в возрасте от 2 месяцев до 7 лет. В случае если в многодетной семье имеется только один или двое дошкольников, организовать семейный детсад можно, пригласив в него дошкольников из других семей.
Постановление Правительства Москвы от 30.10.2007 № 951-ПП
Частный детский сад — это любая форма присмотра за детьми, организованная юрлицом, ИП или самозанятым, то есть не государством. В таком саду за ребенком могут просто присматривать и ухаживать, а могут чему-то обучать по программе, если у сада есть на это лицензия.
Группа по присмотру и уходу — это один из вариантов частного детского сада, который не требует лицензии. В такой группе за детьми присматривают, кормят, гуляют, играют, но не имеют права заниматься их обучением и подготовкой к школе.
Центр развития детей или детский клуб — это тоже варианты частного детского сада, которые могут работать как по лицензии, так и без нее. Такие центры могут создавать самозанятые — при условии, что они не занимаются образовательной деятельностью, ИП, некоммерческие (НКО) и коммерческие организации. Для последних образовательная деятельность, если центр ею занимается, всегда будет не основной, а дополнительной.
В детских клубах и подобных заведениях дети не остаются на весь день, не едят, не спят. Соответственно, к ним не применяются некоторые нормы санпина, например не нужно организовывать спальные места. Но открывать детский сад полного дня под видом центра развития незаконно, даже если у него есть лицензия.
Негосударственная образовательная организация — это детский сад или детский центр, оформленный как некоммерческое юрлицо. Для такого юрлица образовательная деятельность всегда основная. До 2013 года только с такой формой собственности вообще можно было получить лицензию. Сейчас некоммерческие организации, как правило, оформляют те, кто рассчитывают на субсидии от государства и гранты от фондов.
Сейчас некоммерческие организации, как правило, оформляют те, кто рассчитывают на субсидии от государства и гранты от фондов.
Различия детских дошкольных заведений
| Название | В какой форме создается | Нужна ли лицензия | Что могут делать для детей | Что получают от государства |
|---|---|---|---|---|
| Семейная дошкольная группа | Группа при муниципальном садике | Нет. Но создать может только многодетная семья, где есть хотя бы один дошкольник | Присмотр, уход и обучение | Зарплата воспитателю, оплата питания, учебные пособия, возможность приходить на занятия в муниципальный детский сад |
| Частный детский сад | ИП, самозанятость, юрлицо | Да, но доступна только для ИП и юрлиц, если они занимаются образовательной деятельностью | Присмотр, уход, обучение | Субсидии |
| Группа по присмотру и уходу | ИП, самозанятость, юрлицо | Нет | Присмотр и уход | — |
| Центр развития детей, детский клуб | ИП, самозанятость, юрлицо | Да, но доступна только для ИП и юрлиц, если они занимаются образовательной деятельностью | Обучение, присмотр | — |
| Негосударственная образовательная организация | Некоммерческое юрлицо | Да | Присмотр, уход, обучение | Субсидии |
Семейная дошкольная группа
В какой форме создается
Группа при муниципальном садике
Нужна ли лицензия
Нет. Но создать может только многодетная семья, где есть хотя бы один дошкольник
Но создать может только многодетная семья, где есть хотя бы один дошкольник
Что могут делать для детей
Присмотр, уход и обучение
Что получают от государства
Зарплата воспитателю, оплата питания, учебные пособия, возможность приходить на занятия в муниципальный детский сад
Частный детский сад
В какой форме создается
ИП, самозанятость, юрлицо
Нужна ли лицензия
Да, но доступна только для ИП и юрлиц, если они занимаются образовательной деятельностью
Что могут делать для детей
Присмотр, уход, обучение
Что получают от государства
Субсидии
Группа по присмотру и уходу
В какой форме создается
ИП, самозанятость, юрлицо
Нужна ли лицензия
Нет
Что могут делать для детей
Присмотр и уход
Что получают от государства
—
Центр развития детей, детский клуб
В какой форме создается
ИП, самозанятость, юрлицо
Нужна ли лицензия
Да, но доступна только для ИП и юрлиц, если они занимаются образовательной деятельностью
Что могут делать для детей
Обучение, присмотр
Что получают от государства
—
Негосударственная образовательная организация
В какой форме создается
Некоммерческое юрлицо
Нужна ли лицензия
Да
Что могут делать для детей
Присмотр, уход, обучение
Что получают от государства
Субсидии
Как домашние сады регулируются государством
ИП, ООО или некоммерческие организации любого типа в своей работе руководствуются гражданским и трудовым кодексами, законом «О лицензировании отдельных видов деятельности» и Положением о лицензировании образовательной деятельности.
В семейных дошкольных группах многодетный родитель становится воспитателем или помощником воспитателя в одном из государственных детских садов, а не предпринимателем. Его оформляют по трудовому кодексу. Подробно деятельность домашних и семейных групп регулируется Письмом Минобрнауки от 22.02.2017 № 08-364 и региональными нормативными актами. Например:
Например, по требованиям санпинов если дети проводят в частном садике меньше четырех часов, то не нужны кровати и место для приема пищи, а если больше — придется оборудовать кухню и спальню. Если садик занимается только присмотром и уходом, то не должно быть обучающих занятий. Если обучение проводится и есть лицензия, то должна быть образовательная программа, а воспитатели должны иметь педагогическое образование.
С 2013 года санпины смягчились, например частные детские сады разрешили открывать в жилых помещениях, а комплектность групп стали определять не по количеству детей, а по количеству квадратных метров на ребенка.
А с января 2021 года в силу вступили новые санитарные правила, которые, напротив, усложнили жизнь детским учреждениям. Например, по новым требованиям любая дошкольная организация теперь обязана иметь собственную территорию для прогулок детей. Исключение — группы по присмотру и уходу, которые не занимаются образованием, — они могут использовать детские игровые площадки во дворах жилых домов.
п. 3.1.2, абз. 8 п. 3.1.11 СП 2.4.3648-20
Плюсы и минусы бизнеса
Если вы не подходите под условия создания семейной дошкольной группы, то свой детский сад становится своим бизнесом, в котором есть плюсы и минусы.
Плюсы:
- ваши дети находятся под вашим же присмотром и вы уверены, что их хорошо кормят и не обижают;
- это дополнительный доход для родителей.
Минусы:
- если детский сад организован в вашей квартире, придется часть ее выделить для работы — ваше личное пространство уменьшится;
- детский сад поблизости может не понравиться соседям — они начнут мучить вас жалобами и проверками.

Не стоит также забывать, что для организации детского сада, даже маленького и без лицензии, нужно официально регистрироваться, платить налоги, вкладываться в качественное оборудование. Покупать подходящие по санпинам стройматериалы для ремонта и игрушки. То есть соответствовать всем требованиям законов.
Риски и проблемы при открытии домашнего детского сада. Главная проблема при организации детского учреждения — это помещение. Детский сад может располагаться и в жилом, и в нежилом фонде, но в любом случае помещение должно соответствовать всем нормативам — подробно о них ниже.
Прекрасно, если для детского сада у вас есть просторная квартира, желательно на нижнем этаже. При этом запрещено размещать любые детские учреждения в цокольном этаже. Еще лучше, если вы владеете коттеджем или частным домом. От размера и обустроенности помещения зависит то, сколько детей вы сможете принимать, а также сможете ли вы получить лицензию. Если подходящей недвижимости нет, то помещение придется арендовать.
Например, у меня частный дом, и я могу выделить 30 м² под игровую комнату. По санитарным правилам с такой площадью я смогу принять до 12 детей до 3 лет — по 2,5 м² на ребенка, или 15 детей старше 3 лет — по 2 м² на ребенка.
п. 3.1.1 СП 2.4.3648-20
Если ваш детский сад располагается в квартире, нужно учесть интересы других жильцов многоквартирного дома. Соседей могут не устроить шум и крики, незнакомые люди — родители детей, которые по утрам и вечерам заходят в подъезд, игры детей на придомовой территории, которая входит в общую собственность жильцов и т. д. Особенно внимательно нужно относиться к соседям, если вы арендуете помещение. В этом случае лучше заранее договориться с собственником о том, что в арендуемой недвижимости будет детский сад.
Решение Набережночелнинского городского суда № 2-4129/2018 от 03.05.2018 по иску о запрете размещения дошкольной группы по присмотру и уходу за детьми в квартире
Еще одна проблема — лицензирование. По закону, если вы не оказываете образовательных услуг, лицензия вам не нужна. Но в этом случае нужно внимательно относиться к собственной рекламе — нельзя упоминать в ней слова «образование», «подготовка к школе», и даже название «детский сад» может вызвать подозрение.
Но в этом случае нужно внимательно относиться к собственной рекламе — нельзя упоминать в ней слова «образование», «подготовка к школе», и даже название «детский сад» может вызвать подозрение.
Чтобы оформить лицензию, нужно набрать персонал, подготовить помещение и получить разрешения Роспотребнадзора о соответствии помещения нормативам и Рособрнадзора о соответствии образовательной программы. О порядке лицензирования расскажу дальше.
Открытие полноценного детского сада требует вложений, которые могут не окупиться. Особенно если вы арендуете помещение, а не работаете в своей квартире или доме. В больших городах бизнесу будет мешать высокая конкуренция, в маленьких — низкий доход потенциальных клиентов.
Форум «Детский центр как бизнес» — ответы на разные вопросы по организации детских садов
Кроме того, все, что связано с детьми, — заведомо опасный бизнес. Дети могут заболеть, упасть, отравиться, подраться, испортить вещи — за все эти неприятности ответственность несет детский сад.
Главное, чтобы были довольны ваши клиенты — дети и их родители
Иван Сорокин
основатель сети детских садов Smile Fish
Начинающий предприниматель, который хочет открыть детский сад, может столкнуться с несколькими трудностями. Чаще всего не удается открыться в установленный срок, вложения превышают запланированные — хотел вложить два миллиона, а вложил уже три, или расходы превышают доходы — время идет, предприниматель платит аренду, а детей все еще недостаточно.
Главное в такой ситуации — как следует взвесить все с самого начала, продолжать работать и оказывать качественную услугу, даже если в первые месяцы нет полного набора и ожидаемых доходов.
Не нужно усложнять себе жизнь и сразу брать на себя слишком много. Вместо садика в 500 м² проще открыть небольшой частный сад на первом этаже жилого дома, не устанавливать космическую цену за свои услуги, а узнать, какая цена адекватна для этого района. Вместо того чтобы тратить усилия и деньги на лицензирование, начните с группы по присмотру и уходу.
Например, у нас 30 садиков в Москве и нет образовательной лицензии. Потому что мы не предлагаем образовательные услуги и все занятия проводим только в игровой форме. Наша цель, чтобы ребенку было интересно и комфортно ходить в наши сады, а родители могли спокойно его оставить. С таким форматом гораздо легче работать и легче открыться.
Начинающие предприниматели часто боятся проблем с разными службами, с соседями в домах, где они арендуют помещение. И за этими страхами забывают о главном — о детях. И тут совет один — делайте все по закону, и никакие соседи и проверки вам будут не страшны.
Кто может открыть семейный детский сад
Если речь о семейной дошкольной группе, то организовать ее может только многодетный родитель — неважно, мама или папа. Требования к таким группам зависят от региона. Например, в Москве наполняемость групп должна быть от 3 до 5 детей, а в Тульской области — от 5 до 8 детей. При этом в группе может быть как несколько собственных детей многодетного родителя-воспитателя, так и один.
Частный детский сад может открыть любой человек — достаточно зарегистрировать ИП, ООО или некоммерческую организацию.
При этом важно учитывать, что в сфере образования, воспитания и развития несовершеннолетних не могут работать люди с судимостями, в том числе погашенными, осужденные за преступления против жизни и здоровья, свободы, чести и достоинства личности, за подрыв конституционного строя и некоторые другие. Эту информацию об ИП налоговая обязательно запросит при регистрации.
ст. 325.1 ТК РФ
подп. «к» п. 1 ст. 22.1 закона о государственной регистрации юрлиц и ИП
Открывать юрлицо учредителю с судимостью закон не запрещает. Но такой учредитель все равно не сможет заниматься педагогической деятельностью в своей организации.
Когда нужна лицензия. Для открытия частного детского сада лицензия понадобится во всех случаях, когда в саду оказывают образовательные услуги и есть нанятые преподаватели, обучающая программа, расписание занятий и методические пособия. Также всегда придется лицензировать деятельность некоммерческим образовательным организациям.
Также всегда придется лицензировать деятельность некоммерческим образовательным организациям.
ст. 91 закона об образовании
Без лицензии работать с детьми можно, только если:
- Частный сад занимается только уходом и присмотром за детьми, в его рекламе и расписании нет занятий, в работе не используются методические пособия, а в штате нет преподавателей.
- Владелец детского сада — ИП, который имеет педагогическое образование и лично оказывает образовательные услуги.
- Это семейная дошкольная группа при государственном детском садике.
Как открыть садик при поддержке государства
Государство приветствует появление частных детских садов и семейных дошкольных групп — есть даже специальный федеральный проект «Содействие занятости» в рамках национального проекта «Демография». В нем прописано, что государство должно способствовать появлению мест в детских садах, в том числе и поддерживать частные дошкольные учреждения. Но чтобы воспользоваться льготами, программами и субсидиями, нужно делать все строго по закону.
Проще всего сотрудничать с государством многодетным мамам, которые решили оформить семейную группу. Приведу пример, как они работают.
Наталья много лет работает воспитателем детского сада на дому. Сначала она занималась со своими детьми, а когда кто-то вырастал, брала в группу приходящих малышей. Все это время она оформлена воспитателем в государственном образовательном комплексе в Москве. Она получает зарплату воспитателя и деньги на питание из расчета 133,05 Р в день на дошкольника.
Количество семейных дошкольных групп в Москве
| Год | Количество семейных групп | Количество воспитанников |
|---|---|---|
| 2008 | 200 | 650 |
| 2010 | 426 | 1300 |
| 2015 | 657 | 2300 |
| 2019 | 979 | 3200 |
Количество семейных групп
200
Количество воспитанников
650
Количество семейных групп
426
Количество воспитанников
1300
Количество семейных групп
657
Количество воспитанников
2300
Количество семейных групп
979
Количество воспитанников
3200
Источник: semds. ru
ru
Детские сады, оформленные как некоммерческие организации, а также с недавних пор ИП и ООО, могут рассчитывать на государственную помощь с помещением, а также дополнительные субсидии и специальные программы. Эти программы разные в разных регионах.
Например, в Воронеже или Новгородской области государство оплачивает часть стоимости обучения частным детским садам, когда они берут детей из муниципальной очереди. Выплачивается та же сумма, которую платят на одного воспитанника государственным садам, — примерно 20 000 Р в месяц в зависимости от региона. А родителям остается только оплатить разницу.
Поручение Правительства РФ «О мерах по развитию частных дошкольных образовательных организаций»
Чтобы попасть в частный садик по такой программе, родители, вставая в очередь, выбирают его из специального списка. Чтобы садику попасть в этот список и получить финансирование, нужно обратиться в региональный департамент образования.
Оформление документов и получение лицензии
Проще всего — открыть семейную дошкольную группу, сложнее — коммерческую или некоммерческую организацию. Расскажу подробно, как оформить документы и для первого, и для второго вариантов.
Расскажу подробно, как оформить документы и для первого, и для второго вариантов.
Чтобы открыть семейную дошкольную группу, многодетный родитель выбирает учебное заведение своего региона, которое уже поддерживает или готово поддержать программу развития семейных детских садов. На этапе подготовки он также должен привести помещение, в котором предполагается семейный детский сад, в соответствие с санитарными нормами и найти нужное количество воспитанников, если собственных дошкольников в семье не хватает.
Затем родитель подает два заявления — об открытии семейного детского сада и о принятии его на работу на должность воспитателя. Образовательное учреждение проводит проверку жилищно-бытовых условий и заключает договоры с родителем — трудовой и об открытии семейной дошкольной группы.
Для такой формы детского сада не нужно открывать отдельный счет, выбирать систему налогообложения или платить взносы в налоговую и ФСС — этим занимается организация-работодатель.
Регистрация частного детского садика возможна в любой правовой форме — юрлицо, ИП или самозанятость.
Самозанятые не могут заниматься образовательной деятельностью, потому что прямо не названы в статье 21 закона об образовании. Но они могут организовывать группы по присмотру и уходу за детьми — для этого даже не нужна лицензия. И естественно, в этом случае нельзя нанимать сотрудников.
Еще один простой вариант — регистрация в качестве ИП. Мы уже писали подробную статью о том, как открыть ИП: сделать это можно в том числе через портал госуслуг.
Для открытия частного детского сада ИП нужно выбрать ОКВЭД:
- если у ИП нет педобразования — 88.91 «Присмотр и уход за детьми»;
- если есть педобразование или ИП планирует получать лицензию — любой из ОКВЭДов, связанных с образованием. Например, 85.11 «Образование дошкольное» или 85.41.9 «Образование дополнительное детей и взрослых прочее, не включенное в другие группировки».

Ну и наконец, частный детский сад может быть некоммерческой организацией — подойдет форма частного учреждения или автономной некоммерческой организации. Что такое некоммерческие организации, зачем они нужны, как их регистрировать и где они могут брать деньги, рассказали в отдельной статье.
Открытие расчетного счета для детского сада происходит по стандартной процедуре: нужно обратиться в любой банк с заявлением и документами о компании. Как правило, счет открывается после регистрации бизнеса. ИП могут работать и без него, но это сложнее: есть ограничения по расчетам наличными. Самозанятым, как правило, достаточно своего личного банковского счета — отдельный бизнес-счет не требуется.
Система налогообложения. Частному детскому садику, как и любому другому бизнесу, нужно выбрать подходящую систему налогообложения. Для детского сада подходят УСН «Доходы», УСН «Доходы минус расходы» или патент.
Самозанятые работают на специальном налоговом режиме — платят налог на профессиональный доход.
Постановка на учет в ФСС, ПФР как отдельное действие не требуется — налоговая сама направляет туда информацию о регистрации юрлица или ИП. Исключение: если у ИП появляются сотрудники, ему нужно направить в ФСС заявление о регистрации в качестве страхователя и некоторые другие документы.
Регистрация и снятие с учета в ПФР
Правила регистрации в ФСС
Самозанятые становятся на учет в ПФР по своему желанию — если хотят делать отчисления на будущую пенсию.
Согласование деятельности с департаментом образования — это и есть лицензирование. Никакие другие согласования вне процесса лицензирования для открытия частного детского сада не нужны.
Заявление на получение лицензии можно подать через портал госуслуг, срок его рассмотрения — до 45 дней. Стоимость лицензии — 7500 Р. Лицензия действует бессрочно.
Перед тем как обращаться за лицензией, нужно:
- Подготовить помещение, соответствующее санпинам.
- Нанять преподавателей.
- Написать образовательную программу.
- Получить положительные заключения от Рособрнадзора, который проверит преподавателей, учебные пособия и образовательную программу.
- Получить положительные заключения от Роспотребнадзора — он проверяет соответствие помещения санпинам, наличие договоров с поставщиками продуктов и на вывоз мусора. А для НКО — еще и пожарного надзора.

Для организации садика в квартире или доме договоры с пожнадзором необязательны: он приходит уже после открытия и не нужен для получения лицензии. Но их все равно могут спросить. Пожарные требования зависят от количества детей в группе и типа помещения — отдельно стоящее или часть многоквартирного дома. Например, если в садике до 10 детей, то пожарные нормы для него такие же, как и для обычной квартиры. А если детей больше, то будут дополнительные требования, например возможность организовать запасной выход.
Франшиза. Еще один способ открыть домашний детский сад — купить франшизу.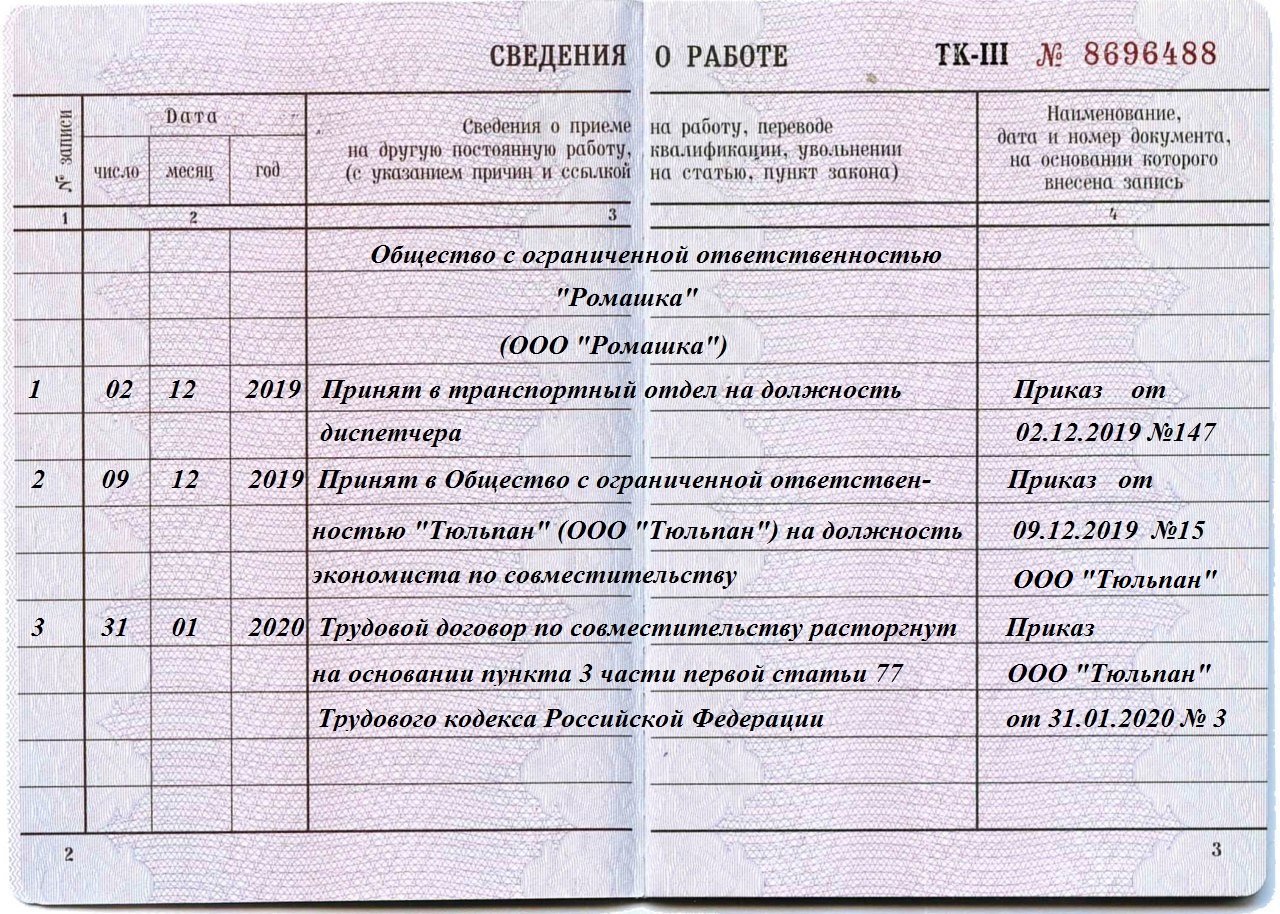 По франшизе можно открыть любой вариант домашнего садика, кроме семейной группы.
По франшизе можно открыть любой вариант домашнего садика, кроме семейной группы.
При таком подходе вам придется заплатить паушальный взнос, а еще выплачивать роялти, то есть отдавать процент или фиксированную сумму из своего дохода. Зато при покупке франшизы предпринимателю не нужно думать об оформлении и дизайне, покупке оборудования, методических пособиях, разработке образовательных программ. Обычно в стоимость франшизы также входит помощь юристов, рекламная продукция, фирменный стиль и помощь в обучении персонала. Но при этом нельзя будет вносить в работу вашего будущего садика никаких изменений.
Чтобы лучше разобраться с тем, как вести бизнес по франшизе и не прогореть, посмотрите нашу подборку статей.
Что нужно для открытия детского сада в своей квартире
Требования к помещению, оборудованию, персоналу и питанию установлены правилами — СП 4.13130.2013 и СП 2.4.3648-20 и санпинами — 2.3/2.4.3590-20 и 1.2.3685-21. За образовательную часть отвечает Рособрнадзор.
Требования к помещению. Организовать частный сад можно как в жилом, так и нежилом помещении. К последним предъявляется намного больше требований. А семейную детскую группу можно открыть только в жилом помещении.
При размещении детей в квартире или доме учитывают общую площадь помещения, наличие водоснабжения, канализации, количество света, качество отделки, этажность и наличие запасных выходов.
Требования к жилым помещениям частных детских садов — по СП 2.4.3648-20
| Условие | Требования | Пункт СП |
|---|---|---|
| Количество помещений | Нужны: — место для хранения верхней одежды и обуви; — игровая комната; — место для сна; — кухня и место, где дети могут кушать, если они находятся в садике больше 4 часов; — место, где дети могут пить; — место для хранения белья, если дети спят в садике; — шкаф для хранения уборочного инвентаря; — туалет и умывальная комната — их можно совместить в одном помещении | 3. 1.11 1.11 |
| Площадь помещения | В игровой комнате без учета мебели: — для детей до 3 лет — не менее 2,5 м² на ребенка; — для детей от 3 до 7 лет — не менее 2 м² на ребенка. В спальной комнате: | 3.1.1 |
| Количество света | Уровень искусственной освещенности для детей дошкольного возраста в игровых комнатах — не менее 400 люкс. В одном помещении не должно быть разных типов ламп или ламп с разным светоизлучением | 2.8.5 |
| Качество отделки | Все строительные и отделочные материалы должны быть лицензированы и безопасны для человека — это должно подтверждаться документами. Нужно использовать только отделку и материалы, которые можно помыть. Полы, стены и потолки должны быть без дефектов, грибковых поражений, следов протеканий | 2.5.1—2.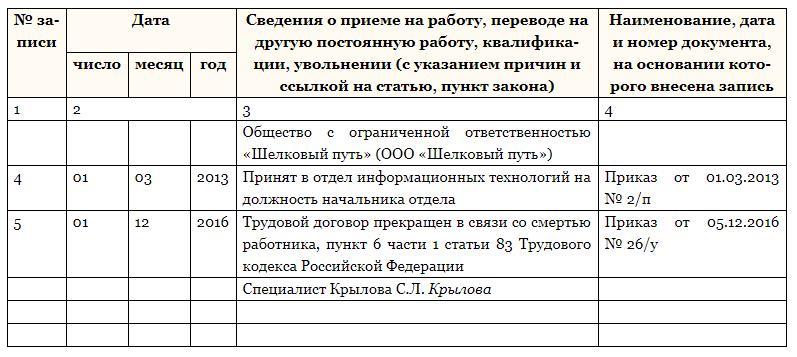 5.3 5.3 |
| Этажность | Помещения для детей нельзя размещать на цокольных и подвальных этажах — их можно использовать только для хозяйственных нужд | абз. 8—9 п. 2.3.1 |
Количество помещений
Требования
Нужны:
— место для хранения верхней одежды и обуви;
— игровая комната;
— место для сна;
— кухня и место, где дети могут кушать, если они находятся в садике больше 4 часов;
— место, где дети могут пить;
— место для хранения белья, если дети спят в садике;
— шкаф для хранения уборочного инвентаря;
— туалет и умывальная комната — их можно совместить в одном помещении
Площадь помещения
Требования
В игровой комнате без учета мебели:
— для детей до 3 лет — не менее 2,5 м² на ребенка;
— для детей от 3 до 7 лет — не менее 2 м² на ребенка. В спальной комнате:
— для детей до 3 лет — не менее 1,8 м² на ребенка;
— для детей от 3 до 7 лет — не менее 2,0 м² на ребенка
Количество света
Требования
Уровень искусственной освещенности для детей дошкольного возраста в игровых комнатах — не менее 400 люкс.
В одном помещении не должно быть разных типов ламп или ламп с разным светоизлучением
Качество отделки
Требования
Все строительные и отделочные материалы должны быть лицензированы и безопасны для человека — это должно подтверждаться документами.Нужно использовать только отделку и материалы, которые можно помыть. Полы, стены и потолки должны быть без дефектов, грибковых поражений, следов протеканий
Пункт СП
2.5.1—2.5.3
Требования
Помещения для детей нельзя размещать на цокольных и подвальных этажах — их можно использовать только для хозяйственных нужд
Пункт СП
абз. 8—9 п. 2.3.1
Обстановка помещения детского сада на дому. В первую очередь для детей потребуется мебель. Если это полноценный детский сад, то нужны столы, стулья и кроватки, которые должны соответствовать росту и возрасту детей.
п. 2.4.3, абз. 3, 5 п. 3.1.11, п. 3.2.3 СП 2.4.3648-20
Также на каждого ребенка требуются отдельное полотенце, сиденья на унитаз или горшок и комплект постельного белья. А игрушки используются только сделанные из безопасных материалов — пластиков, разрешенных к использованию в быту, дерева, резины — и только те, которые можно помыть.
А игрушки используются только сделанные из безопасных материалов — пластиков, разрешенных к использованию в быту, дерева, резины — и только те, которые можно помыть.
Организация питания. Кормить детей можно едой собственного приготовления или готовой на заказ. В любом случае на кухне детского садика должны быть отдельные холодильники или полки для разных продуктов и посуды, а также все необходимое технологическое и моечное оборудование, инвентарь и посуда.
п. 8.6—8.6.8 Санпина 2.3/2.4.3590-20
Кроме того, все нужно подписывать: посуду и полки — в зависимости от того, какие продукты в них готовятся или хранятся, продукты — в зависимости от того, где и когда они были куплены. Также нужно вести журнал бракеража готовой продукции — оставлять понемногу еды для проверки на случай, если у кого-то из детей будут проблемы со здоровьем.
Санпин также регламентирует меню для детей, количество приемов пищи, продукты, которые можно и нельзя есть детям. Например, детям нельзя есть пирожные и торты с кремом, макароны по-флотски с фаршем, грибы, чипсы и даже яичницу-глазунью.
Например, детям нельзя есть пирожные и торты с кремом, макароны по-флотски с фаршем, грибы, чипсы и даже яичницу-глазунью.
Полный перечень продукции, которая не допускается при организации питания детей, в Приложении № 6 к Санпину 2.3/2.4.3590-20
Кроме того, в садике обязательно должно быть организовано место, где дети смогут попить воду — требования к нему установлены в п. 8.4—8.4.6 СП 2.4.3648-20.
Разработка методики и обучающей программы раннего развития. Если частный детсад не заявлен как образовательное учреждение, а занимается только присмотром и уходом, то обучающая программа ему не нужна. В противном случае без полноценной программы не получить лицензию на образовательную деятельность.
Для получения лицензии программа должна соответствовать требованиям федерального государственного образовательного стандарта дошкольного образования — ФГОС и Примерной основной образовательной программе дошкольного образования.
Самый простой вариант — использовать готовые программы, одобренные для дошкольников. Их можно найти в интернете, например на сайте Федерального института развития образования, или купить у уже действующих сетевых детских садов — некоторые из них предлагают такую услугу.
Если вы педагог, то можете самостоятельно подготовить программу, опираясь, например, на программы, которыми пользуются в муниципальных детских садах, или популярные методики развития детей — Зайцева, Монтессори, Вальдорфскую и другие.
Семейным группам, которые организуют многодетные родители, программу обучения, а также некоторые пособия предоставляет образовательная организация, к которой они прикреплены. При желании ребенок может обучаться в семейной группе и ходить в государственный детский сад на отдельные занятия, например музыку или физкультуру.
Наем персонала в случае с частными детскими садами зависит от формы бизнеса: самозанятые не могут никого нанимать по трудовым договорам, ИП, ООО и НКО — могут, но с условиями.
Для подбора персонала нужно оформить штатное расписание, определиться с численностью работников, специалистами, которые потребуются для реализации образовательных программ и другой деятельности детского сада.
У педагогических работников обязательно должно быть высшее или среднее профессиональное образование. Кроме того, все сотрудники детсада при приеме на работу обязаны пройти медицинский осмотр.
ст. 46 закона об образовании
ст. 213 ТК РФ
Если у вас лицензированный детский сад, то воспитатели, которые у вас работают, пользуются всеми правами и льготами, которые есть у воспитателей в обычном детском саду, например им засчитывается педагогический стаж.
Выбор воспитателя и требования к его образованию. Работать воспитателем, в соответствии со статьей 46 закона об образовании, могут только люди, получившие или получающие среднее специальное или высшее педагогическое образование. Если такого образования нет, но есть другое высшее образование, будущий воспитатель может пройти курсы повышения квалификации по специальности «Воспитатель ДОУ».
Если в детском саду планируется использовать какую-то конкретную методику, воспитатель должен пройти курсы подготовки именно по этому направлению. Например, в вальдорфских школах есть специальные курсы для педагогов.
В семейных дошкольных группах ситуация иная. Если у родителя нет педагогического образования, его оформляют в ДОУ, к которому прикреплена группа, не воспитателем, а помощником воспитателя. Реальных функций родителя-воспитателя это не меняет, но зарплата в этом случае меньше и педагогический стаж не идет. Для того чтобы родителю стать воспитателем, нужно пройти курсы повышения квалификации или отучиться в колледже или вузе.
Кроме образования от воспитателей еще требуется отсутствие судимостей и опасных болезней, при которых нельзя работать с детьми, например туберкулеза в открытой форме, алкоголизма и наркомании.
Искать педагогов можно через сайты по поиску работы, например на «Авито» или «Хедхантере». А еще в педагогических вузах и колледжах — там можно разместить объявления.
Нет единого мнения, какие воспитатели лучше — начинающие или с опытом. Первые часто ближе к детям по духу и не испорчены штампами, а вторые — лучше смогут организовать учебу и справятся с нештатными ситуациями. В любом случае нужно смотреть на конкретного человека и выбирать того педагога, которому вы доверили бы своего ребенка.
Набор детей. В семейной группе в зависимости от региона может быть от 3 до 8 детей, в частном садике ограничения диктуются только площадью помещения — на одного ребенка должно быть 2—2,5 м². Обычно всех детей в такую группу ищут среди знакомых или родственников или в группах единомышленников, которые предпочитают домашнее образование.
Найти детей в частный детский сад сложнее: нужно убедить родителей, что в таком саду лучше, чем в государственном, а в вашем частном саду лучше, чем у конкурентов. Для этого не только нужно постоянно давать рекламу, но и действительно поддерживать качество образования, чистоту и внимательно следить за работой воспитателей. Если услуги будут на высоком уровне, родители приведут своих знакомых.
Реклама и привлечение клиентов. Поначалу группы в небольших домашних садиках набираются благодаря сарафанному радио из знакомых и их знакомых. Но если нужно набрать 10—15 детей, которые будут регулярно посещать садик и вовремя оплачивать услуги, то придется заняться рекламой. Здесь есть несколько вариантов: реклама в соцсетях, печатные объявления и визитки, реклама в СМИ.
Чтобы искать клиентов в соцсетях, создайте сообщество или страницу своего детского сада и разместите рекламу в родительских группах или на тематических форумах.
Родительские форумы:
forum.littleone.ru — в Петербурге,
lubopyshka.ru — в Ярославле,
«Мамы района Ясенево и ЮЗАО» — в Москве
Печатные объявления можно разместить на досках объявлений, на остановках и у подъездов. Визитки — оставить в детских учреждениях: домах культуры, библиотеках, поликлиниках, роддомах.
Реклама в СМИ больше подойдет большим организациям, а не крошечным садикам на 10—15 детей, но если вы в небольшом городе, то можно попробовать дать объявление в местную газету или на радио.
Чтобы лучше разобраться в продажах и привлечении клиентов, почитайте нашу подробную статью с основами маркетинга для бизнеса.
В любом случае рекламировать нужно только реальные услуги и только ту деятельность, которой вы имеете право заниматься. Если у вас нет лицензии на образовательную деятельность, то в рекламе нельзя рассказывать о подготовке к школе и углубленном английском — говорите, например, о группах по присмотру и уходу.
Договоры с родителями заключать обязательно. Прописывайте в них права и обязанности родителей и руководства детского садика, а также уточняйте размер оплаты и период, за который платят родители, — за месяц или по часам. Важно также прописать, как будут считаться и оплачиваться дни, в которые ребенок не посещал детский сад. Например, большинство частных садов не возвращает деньги за дни, пропущенные по болезни, и родители заранее соглашаются на такие условия, если хотят попасть в хороший детский сад.
Частный детский сад может разработать собственную форму договора или взять готовую из интернета.
Расходы, прибыльность и сроки окупаемости бизнеса
В каждом конкретном случае расходы, прибыльность и окупаемость бизнеса будут свои. Эти параметры зависят от множества факторов: выбираете вы для бизнеса ИП или ООО, открываете сад в своей квартире или в арендованном нежилом помещении, где оно находится, нужно ли там делать ремонт и закупать специальное оборудование, сколько у вас воспитателей, педагогов и других сотрудников, как они оформлены и т. д. Поэтому перед началом любого проекта рекомендую составлять собственный бизнес-план.
Для примера я рассчитаю расходы и доходы от открытия частного детского сада в моем доме.
Выбор формы бизнеса. Если бы я открывала детский сад, я бы регистрировалась в качестве ИП: я уже была предпринимателем и мне знакома эта форма деятельности. Кроме того, ИП проще открыть и закрыть, чем ООО.
Но есть и минус — если я как ИП залезу в долги и мой бизнес прогорит, то я буду отвечать своим имуществом. Образование я бы сделала дополнительным видом деятельности.
Стоимость регистрации в качестве ИП — 0 Р, если подавать заявление онлайн. Если регистрироваться офлайн — 800 Р.
Открытие расчетного счета для ИП в большинстве банков бесплатно. Обслуживание обходится в зависимости от выбранного тарифа от 500 до 4000 Р в месяц.
Работать я бы предпочла по упрощенной системе налогообложения — УСН 6%. Я и раньше работала по этой системе, она подходит тем, кто оказывает услуги и не имеет высоких текущих расходов и доходов. Для организации частного детского сада ИП также может быть выгодно оформить патент, его стоимость в регионах зависит от среднего дохода в отрасли и часто ниже стандартных налоговых режимов.
Рассчитать стоимость патента можно на сайте ФНС
Помещение арендовать мне не нужно: у меня есть двухэтажный загородный дом, часть первого этажа — 50 м² — я могу задействовать для детского сада. По санпину на этой площади я могу разместить 25 детей от 3 до 7 лет, но я вряд ли справлюсь больше чем с 15.
Я не буду тратиться на аренду, но мне понадобятся деньги, чтобы отремонтировать комнаты, которые я отведу под детский сад, — обшить и покрасить стены.
Примерная стоимость ремонта для моего дома 200 000 Р.
Из оборудования мне понадобятся посуда, горшки, игрушки. А еще придется купить детскую мебель и различную технику.
Примерная стоимость оборудования — 690 000 Р
| Детская мебель — столы стулья, кровати, шкафчики | 300 000 Р |
| Техника — стиральная машина, холодильник, компьютер, аудиосистема, проектор | 150 000 Р |
| Текстиль — постельное белье, полотенца | 80 000 Р |
| Игрушки, игровые зоны и развивающие игры | 60 000 Р |
| Игрушки и игровая площадка для улицы | 50 000 Р |
| Спортивный комплекс и спортивные снаряды | 30 000 Р |
| Посуда | 15 000 Р |
| Горшки и сиденья на унитаз | 5 000 Р |
Детская мебель — столы стулья, кровати, шкафчики
300 000 Р
Техника — стиральная машина, холодильник, компьютер, аудиосистема, проектор
150 000 Р
Текстиль — постельное белье, полотенца
80 000 Р
Игрушки, игровые зоны и развивающие игры
60 000 Р
Игрушки и игровая площадка для улицы
50 000 Р
Спортивный комплекс и спортивные снаряды
30 000 Р
Горшки и сиденья на унитаз
5 000 Р
Питание. О питании я бы постаралась договориться с соседней школой — у них есть собственная столовая, которая соответствует требованиям санпина. В этом случае мне нужно будет только договориться о доставке и оборудовать место для приема пищи и хранения продуктов. Стоимость питания на одного ребенка я бы заложила по стоимости в государственном детском саду — 130 Р в день.
Лицензирование. Лицензия обойдется мне в 7500 Р, если я буду оформлять документы сама, и до 35 000 Р, если обращусь к фирме-посреднику. Я, скорее всего, выберу первый вариант.
Кроме того, нужно учесть дополнительные расходы, например на экспертизы и пробы воды и освещения или договоры на дезинфекцию. На эти цели я бы отложила 15 000 Р.
Обучающая программа. Я бы не стала слишком задумываться об образовательной программе в первое время и взяла бы для получения лицензии одну из готовых программ. А к ней бы постепенно добавляла элементы вальдорфской методики и дополнительные занятия.
Сотрудники. Найти хороших сотрудников — самое сложное. Мне обязательно понадобится уборщица и второй воспитатель. Для начала я бы попробовала поискать сотрудников среди своих знакомых. Нужно не забывать, что и за себя, и за сотрудников придется платить налоги и взносы. Также обычно работодатель берет на себя оформление санитарных книжек для сотрудников, это примерно 3500 Р за человека. И нужно учесть зарплаты сотрудников, у нас это около 20 000 Р в месяц.
Договор. Я посоветовалась с экспертом, и он порекомендовал мне не пользоваться типовым договором, а обратиться к юридической компании для составления грамотного соглашения с родителями или купить шаблон договора у любой сети детских садов, которая работает на рынке более пяти лет. Оказывается, такая практика есть. Такой подход поможет избежать многих подводных камней при работе с клиентами и проверках надзорных органов. Договор может обойтись примерно в 10 000 Р.
Реклама. Я бы дала рекламу в рыбинской группе во «Вконтакте» «От мамы к маме» и создала бы группы детского сада в соцсетях. У нас небольшой город, и конкуренция не высокая. На рекламу в первое время я бы выделила не больше 10 000 Р в месяц.
Ежемесячные траты на домашний детский сад — 122 350 Р
| Расходы на питание детей | 130 Р в день на ребенка × 23 рабочих дня = 2990 Р 2990 Р × 15 детей = 44 850 Р |
| Налоги и взносы | 25 000 Р |
| Зарплата второго воспитателя | 20 000 Р |
| Зарплата уборщицы | 12 000 Р |
| Реклама | 10 000 Р |
| Коммунальные платежи | 5000 Р |
| Оплата бухгалтера | 3000 Р |
| Расходы на ведение банковского счета | 1500 Р |
| Бытовая химия и средства гигиены | 1000 Р |
Расходы на питание детей
130 Р в день на ребенка × 23 рабочих дня = 2990 Р2990 Р × 15 детей = 44 850 Р
Налоги и взносы
25 000 Р
Зарплата второго воспитателя
20 000 Р
Зарплата уборщицы
12 000 Р
Реклама
10 000 Р
Коммунальные платежи
5000 Р
Оплата бухгалтера
3000 Р
Расходы на ведение банковского счета
1500 Р
Бытовая химия и средства гигиены
1000 Р
Калькулятор НДФЛ и взносов за работников
Доход. Плата за детский сад в Рыбинске — примерно 15 000 Р в месяц. Как правило, частные садики не возвращают оплату, даже если ребенок болел и тем более если он пропустил день без причины. Если в группе будет 15 детей, то доход составит 225 000 Р в месяц.
Прибыль. При условии, что все 15 детей будут каждый день ходить на занятия, а в хозяйстве не возникнет непредвиденных расходов, прибыль в месяц составит: 225 000 Р − 122 350 Р = 102 650 Р.
Кажется, что это значительная сумма и при таком доходе можно окупить вложения примерно за год. Но это идеальный расклад. В процессе могут появиться непредвиденные расходы, дети могут болеть или переходить в другие садики — это уменьшает ежемесячный доход. Выходит из строя и ломается техника, сантехника, электричество. Кроме того, если у вас нет своего помещения, в расходах придется учитывать арендную плату — еще 25 000—35 000 Р.
Выгодно ли открывать домашний детский сад
- Домашний детский сад — это любая форма присмотра за детьми в домашней обстановке.
- По закону домашний детский сад можно организовать в форме семейной дошкольной группы, прикрепленной к государственному учебному заведению, или частного детского учреждения — образовательного или по присмотру и уходу.
- Относительно выгодно открывать семейную дошкольную группу — работать на государственное образовательное учреждение и получать за это зарплату.
- Для организации частного детского сада, даже маленького и без лицензии, нужно официально регистрироваться, платить налоги, вкладываться в оборудование, ремонт и игрушки, чтобы все соответствовало требованиям законов.
- При определенных условиях и частный детский садик может быть выгодным, но нужно учитывать большие затраты на старте и неизбежные непредвиденные расходы в процессе.
Мидостаурин добавлен к химиотерапии и продолженной монотерапии при остром миелоидном лейкозе с FLT3-ITD | Кровь
Вклад: R.F.S. и H.D. предоставленная концепция и дизайн; RFS, Д. Вебер, WF, HRS, GW, HS, TS, TK, ML, Д. Вольф, JW, DK, KSG, H.-AH, JK, M. Girschikofsky., MR, TS, GH, H. -GD, RS, RG, M. Grießhammer, EL, A. Burchardt, UM, BH, LM, MH, FT, VIG, WH, A. Benner, K.Д., А.Г., П.П. и Х.Д. обеспечили предоставление учебных материалов или пациентов; Р.Ф.С. а Д. Вебер обеспечил сбор и сбор данных; R.F.S., H.D., J.K. и A. Benner предоставили анализ и интерпретацию данных; Р.Ф.С. и H.D. при условии написания рукописи; и RFS, D. Weber, WF, HRS, GW, HS, TS, TK, ML, D. Wolf, JW, DK, KSG, H.-AH, JK, M. Girschikofsky, MR, TS, GH, H. -GD, RS, RG, M. Grießhammer, EL, A. Burchardt, UM, BH, LM, MH, FT, VIG, W.Х., Дж. К., А. Беннер, К. Д., А. Г., П. П. и Х. Д. при условии окончательного утверждения рукописи.
Раскрытие информации о конфликте интересов: R.F.S., K.D. и H.D. получил финансирование исследований от Novartis; R.F.S., K.D., H.D., K.S.G. и D. Wolf получили гонорары от Novartis; Р.Ф.С., К.Д., П.П. и Х.Д. участвовал в консультативных советах Novartis. Остальные авторы заявляют об отсутствии конкурирующих финансовых интересов.
Текущая принадлежность J.К. – отделение гематологии и онкологии, Городская больница Брауншвейга, Германия.
Текущая принадлежность G.H. это отделение внутренней медицины I, Вестпфальцклиникум, Кайзерслаутерн, Германия.
Текущая принадлежность W.H. – III отделение внутренней медицины Университетской клиники Регенсбурга, Германия.
Список организаций и исследователей немецко-австрийской исследовательской группы по борьбе с отмыванием денег, которые участвовали в этом исследовании, приведен в дополнительном Приложении.
Для корреспонденции: Ричард Ф. Шленк, Национальный центр исследования опухолевых заболеваний, Национальный центр опухолевых заболеваний, Im Neuenheimer Feld 130.3, 69120 Гейдельберг, Германия; электронная почта: [email protected].
Потреблениедиетического магния в национальной выборке взрослых в США | Журнал питания
2″> СУБЪЕКТЫ И МЕТОДЫ
Этот анализ основан на данных NHANES, полученных в 1999–2000 гг. (26). Репрезентативная выборка неинституционализированного гражданского населения США была отобрана с использованием стратифицированного многоступенчатого плана выборки. Обученные интервьюеры, используя компьютерную систему личных интервью, проводили собеседования с участниками дома.Участников попросили посетить мобильный экзаменационный центр, где их попросили заполнить дополнительные анкеты, пройти различные обследования и сдать образец крови. В выборку были включены лица с низкими доходами, лица старше 60 лет, афроамериканцы и американцы мексиканского происхождения. Исследование получило одобрение людей, и участников попросили подписать форму информированного согласия.
В отличие от предыдущих NHANES, в которых все участники опрашивались лично, 24-часовой повторный прием пищи проводился либо лично участникам, посещавшим передвижной экзаменационный центр, либо по телефону через 4–10 дней после медицинского осмотра.В подвыборке первичных единиц выборки участники, которых случайным образом назначили на утреннюю сессию в мобильном экзаменационном центре, были опрошены лично. Тех, кого случайным образом назначили на дневную или вечернюю сессию, назначили на телефонное интервью. Однако некоторые из этих участников прошли собеседование по питанию лично, потому что у них не было телефона или из-за проблем с расписанием. Из 4273 взрослых в возрасте ≥20 лет, которые участвовали в отзыве, 3521 были опрошены лично в мобильном экзаменационном центре, а 752 были опрошены по телефону.Обученные двуязычные интервьюеры проводили интервью с использованием компьютерной системы интервьюирования по питанию на английском и испанском языках. Переводчики помогали участникам, которые говорили на других языках. Различные зонды, такие как списки часто забытых продуктов или напитков и специфические пищевые зонды, использовались для улучшения отчетности участников. Информацию о рационе питания обрабатывали с использованием системы анализа потребления пищевых продуктов Техасского университета и базы данных обследований питательных веществ Министерства сельского хозяйства США за 1994–1998 гг. (27,28).
Мы стратифицировали анализы по возрасту, полу, расе или этнической принадлежности (европеоид, афроамериканец и мексиканец).В анализ были включены только участники в возрасте ≥20 лет, которые посещали мобильный экзаменационный центр. Мы представляем в тексте средние значения или проценты ± 1 сем. Различия в средних или геометрических средних были проверены с помощью тестов t . Линейные тренды были исследованы с использованием ортогональных полиномиальных контрастов. Мы использовали SUDAAN (программное обеспечение для статистического анализа коррелированных данных, Research Triangle Park, NC) для анализа, чтобы учесть сложный дизайн выборки.
Мужчины
. Женщины
. . n . Потребление магния
. n . Потребление магния
. . . Среднее значение 1 . Среднее значение 2 . Медиана 3 . . Среднее значение 1 . Среднее значение 2 . Медиана 3 . мг / сут мг / сут Кавказцы 930 352 ± 6 3263 314 901 901 901 256 ± 7 228 ± 7 237 20–30 лет 137 329 ± 14 290 ± 15 299 203 241 ± 13 1313132 211 213 31–50 лет 267 377 ± 13 337 ± 10 353 295 268 ± 11 239 ± 10 249 305 344 ± 11 315 ± 10 325 266 261 ± 6 238 ± 6 241
≥71 лет 2 21 314 ± 15 267 ± 17 280 219 230 ± 10 205 ± 8 205 Афроамериканцы 371 278 ± 10 * 900 ± 15 * 237 439 202 ± 6 * 173 ± 6 * 177 20–30 лет 59 305 ± 27 22270 73 208 ± 14 179 ± 14 176 31–50 лет 153 284 ± 15 * 218 ± 27 * 901 182 208 ± 10 * 181 ± 8 * 183 51–70 лет 107 245 ± 10 * 200 ± 13 * 213 123 192 ± 9 * 169 ± 7 * 179 ≥71 лет 52 227 ± 23 * 19 * 202 61 161 ± 8 * 116 ± 23 * 144 Мексиканские американцы 518 330 ± 9 7131 * 288 297 631 242 ± 7 213 ± 8 221 20–30 лет 99 336 ± 24 291 ± 16 279 16132 279 169 ± 15 235 ± 14 239 31–50 лет 175 345 ± 11 302 ± 10 * 324 201 241 ± 13 213 ± 12 214 51–70 лет 178 291 ± 12 * 258 ± 12 * 262 201 * 21630 ± 14 185 ± 15 * 201 ≥71 лет 66 251 ± 18 * 222 ± 16 * 223 68 9013 ± 15 177 ± 16 188
305 344 ± 11 315 ± 10 325 266 261 ± 6 238 ± 6 241
| . | Мужчины . | Женщины . | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | |||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | |||||||
| мг / сут | мг / сут | ||||||||||||||
| Кавказцы | 930 | 352 ± 6 | 3263 314 901 901 901 256 ± 7228 ± 7 | 237 | |||||||||||
| 20–30 лет | 137 | 329 ± 14 | 290 ± 15 | 299 | 203 | 241 ± 13 | 1313132 211 213|||||||||
| 31–50 лет | 267 | 377 ± 13 | 337 ± 10 | 353 | 295 | 268 ± 11 | 239 ± 10 | 249 | 344 ± 11 | 315 ± 10 | 325 | 266 | 261 ± 6 | 238 ± 6 | 241 |
| ≥71 лет | 2 21 | 314 ± 15 | 267 ± 17 | 280 | 219 | 230 ± 10 | 205 ± 8 | 205 | |||||||
| Афроамериканцы | 371 | 278 ± 10 * 900 ± 15 * | 237 | 439 | 202 ± 6 * | 173 ± 6 * | 177 | ||||||||
| 20–30 лет | 59 | 305 ± 27 | 22270 | 73 | 208 ± 14 | 179 ± 14 | 176 | ||||||||
| 31–50 лет | 153 | 284 ± 15 * | 218 ± 27 * 901 | 182 | 208 ± 10 * | 181 ± 8 * | 183 | ||||||||
| 51–70 лет | 107 | 245 ± 10 * | 200 ± 13 * | 213 | 123 | 192 ± 9 * | 169 ± 7 * | 179 | |||||||
| ≥71 лет | 52 | 227 ± 23 * | 19 *202 | 61 | 161 ± 8 * | 116 ± 23 * | 144 | ||||||||
| Мексиканские американцы | 518 | 330 ± 9 | 7131 * 288297 | 631 | 242 ± 7 | 213 ± 8 | 221 | ||||||||
| 20–30 лет | 99 | 336 ± 24 | 291 ± 16 | 279 16132 | 279 169 ± 15 | 235 ± 14 | 239 | ||||||||
| 31–50 лет | 175 | 345 ± 11 | 302 ± 10 * | 324 | 201 | 241 ± 13 | 213 ± 12 | 214 | |||||||
| 51–70 лет | 178 | 291 ± 12 * | 258 ± 12 * | 262 | 201 | * 21630 ± 14185 ± 15 * | 201 | ||||||||
| ≥71 лет | 66 | 251 ± 18 * | 222 ± 16 * | 223 | 68 | 9013 ± 15 177 ± 16 | 188 | ||||||||
Не скорректированное потребление магния с пищей, Национальное обследование здоровья и питания, 1999–2000 гг.
| . | Мужчины . | Женщины . | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | |||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | |||||||
| мг / сут | мг / сут | ||||||||||||||
| Кавказцы | 930 | 352 ± 6 | 3263 314 901 901 901 256 ± 7228 ± 7 | 237 | |||||||||||
| 20–30 лет | 137 | 329 ± 14 | 290 ± 15 | 299 | 203 | 241 ± 13 | 1313132 211 213|||||||||
| 31–50 лет | 267 | 377 ± 13 | 337 ± 10 | 353 | 295 | 268 ± 11 | 239 ± 10 | 249 | 344 ± 11 | 315 ± 10 | 325 | 266 | 261 ± 6 | 238 ± 6 | 241 |
| ≥71 лет | 2 21 | 314 ± 15 | 267 ± 17 | 280 | 219 | 230 ± 10 | 205 ± 8 | 205 | |||||||
| Афроамериканцы | 371 | 278 ± 10 * 900 ± 15 * | 237 | 439 | 202 ± 6 * | 173 ± 6 * | 177 | ||||||||
| 20–30 лет | 59 | 305 ± 27 | 22270 | 73 | 208 ± 14 | 179 ± 14 | 176 | ||||||||
| 31–50 лет | 153 | 284 ± 15 * | 218 ± 27 * 901 | 182 | 208 ± 10 * | 181 ± 8 * | 183 | ||||||||
| 51–70 лет | 107 | 245 ± 10 * | 200 ± 13 * | 213 | 123 | 192 ± 9 * | 169 ± 7 * | 179 | |||||||
| ≥71 лет | 52 | 227 ± 23 * | 19 *202 | 61 | 161 ± 8 * | 116 ± 23 * | 144 | ||||||||
| Мексиканские американцы | 518 | 330 ± 9 | 7131 * 288297 | 631 | 242 ± 7 | 213 ± 8 | 221 | ||||||||
| 20–30 лет | 99 | 336 ± 24 | 291 ± 16 | 279 16132 | 279 169 ± 15 | 235 ± 14 | 239 | ||||||||
| 31–50 лет | 175 | 345 ± 11 | 302 ± 10 * | 324 | 201 | 241 ± 13 | 213 ± 12 | 214 | |||||||
| 51–70 лет | 178 | 291 ± 12 * | 258 ± 12 * | 262 | 201 | * 21630 ± 14185 ± 15 * | 201 | ||||||||
| ≥71 лет | 66 | 251 ± 18 * | 222 ± 16 * | 223 | 68 | 9013 ± 15 177 ± 16 | 188 | ||||||||
| . | Мужчины . | Женщины . | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | |||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | . | Среднее значение 1 . | Среднее значение 2 . | Медиана 3 . | |||||||
| мг / сут | мг / сут | ||||||||||||||
| Кавказцы | 930 | 352 ± 6 | 3263 314 901 901 901 256 ± 7228 ± 7 | 237 | |||||||||||
| 20–30 лет | 137 | 329 ± 14 | 290 ± 15 | 299 | 203 | 241 ± 13 | 1313132 211 213|||||||||
| 31–50 лет | 267 | 377 ± 13 | 337 ± 10 | 353 | 295 | 268 ± 11 | 239 ± 10 | 249 | 344 ± 11 | 315 ± 10 | 325 | 266 | 261 ± 6 | 238 ± 6 | 241 |
| ≥71 лет | 2 21 | 314 ± 15 | 267 ± 17 | 280 | 219 | 230 ± 10 | 205 ± 8 | 205 | |||||||
| Афроамериканцы | 371 | 278 ± 10 * 900 ± 15 * | 237 | 439 | 202 ± 6 * | 173 ± 6 * | 177 | ||||||||
| 20–30 лет | 59 | 305 ± 27 | 22270 | 73 | 208 ± 14 | 179 ± 14 | 176 | ||||||||
| 31–50 лет | 153 | 284 ± 15 * | 218 ± 27 * 901 | 182 | 208 ± 10 * | 181 ± 8 * | 183 | ||||||||
| 51–70 лет | 107 | 245 ± 10 * | 200 ± 13 * | 213 | 123 | 192 ± 9 * | 169 ± 7 * | 179 | |||||||
| ≥71 лет | 52 | 227 ± 23 * | 19 *202 | 61 | 161 ± 8 * | 116 ± 23 * | 144 | ||||||||
| Мексиканские американцы | 518 | 330 ± 9 | 7131 * 288297 | 631 | 242 ± 7 | 213 ± 8 | 221 | ||||||||
| 20–30 лет | 99 | 336 ± 24 | 291 ± 16 | 279 16132 | 279 169 ± 15 | 235 ± 14 | 239 | ||||||||
| 31–50 лет | 175 | 345 ± 11 | 302 ± 10 * | 324 | 201 | 241 ± 13 | 213 ± 12 | 214 | |||||||
| 51–70 лет | 178 | 291 ± 12 * | 258 ± 12 * | 262 | 201 | * 21630 ± 14185 ± 15 * | 201 | ||||||||
| ≥71 лет | 66 | 251 ± 18 * | 222 ± 16 * | 223 | 68 | 9013 ± 15 177 ± 16 | 188 | ||||||||
Всего 53.0 ± 1,7% участников (мужчины: 48,0 ± 1,9%; женщины: 57,6 ± 2,0%) сообщили об использовании витаминов, минералов или пищевых добавок. Более высокий процент мужчин европеоидной расы (52,7 ± 2,3%) использовали такие добавки, чем мужчины афроамериканского происхождения (29,9 ± 2,8%) и мужчины мексиканского происхождения (26,3 ± 3,4%), а также более высокий процент женщин европеоидной расы (62,3 ± 2,4%). добавки, чем афроамериканки (38,0 ± 2,9%) и мексиканки (39,8 ± 2,2%). Значительно увеличилось употребление с возрастом среди участников европеоидной расы и американцев мексиканского происхождения, но не среди афроамериканцев.
Среди мужчин европеоидной расы и афроамериканцев и женщин европеоидной расы у тех, кто принимал добавки, среднее потребление магния с пищей было значительно выше, чем у тех, кто этого не делал (таблица 2).
ТАБЛИЦА 2Не скорректированное потребление магния с пищей, стратифицированное по статусу пользователя витаминов, минералов или пищевых добавок, Национальное обследование здоровья и питания, 1999–2000 гг.
| . | Пользователи . | Непользователи . | P означает . | P геометрические средства . | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | . | . | |||||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | . | Среднее значение 1 . | Среднее значение 2 . | . | . | |||||||||||
| мг / день | мг / день | ||||||||||||||||||
| Мужчины | 9032 901 | 901 901 | |||||||||||||||||
| Кавказцы | 517 | 368 ± 9 | 331 ± 7 | 412 | 333 ± 9 | 294 ± 9 | 0.015 | 0,001 | |||||||||||
| 20–30 лет | 52 | 320 ± 24 | 287 ± 22 | 85 | 335 ± 19 | 291 ± 21 | 0,625 | 0,905 | 50 лет132 | 391 ± 20 | 350 ± 12 | 135 | 362 ± 15 | 324 ± 14 | 0,229 | 0,160 | |||
| 51–70 лет | 193 36832 | 339 ± 11 | 111 | 297 ± 12 | 272 ± 12 | <0.001 | <0,001 | ||||||||||||
| ≥71 лет | 140 | 342 ± 19 | 302 ± 14 | 81 | 263 ± 19 | 213 ± 30 | 0,003 | 0,015 | 116 | 333 ± 19 | 284 ± 14 | 255 | 255 ± 13 | 199 ± 20 | 0,002 | 0,004 | |||
| 20–30 лет | 16 | 34 279 ± 55 | 43 | 291 ± 27 | 254 ± 22 | 0.462 | 0,659 | ||||||||||||
| 31–50 лет | 47 | 366 ± 30 | 317 ± 24 | 106 | 248 ± 19 | 185 ± 35 | 0,005 | 0,016 | 70 лет37 | 279 ± 21 | 255 ± 17 | 70 | 226 ± 9 | 175 ± 16 | 0,025 | 0,001 | |||
| ≥71 лет | 16 | 172 ± 30 | 36 | 236 ± 30 | 193 ± 24 | 0.379 | 0,600 | ||||||||||||
| Американцы мексиканского происхождения | 151 | 355 ± 16 | 320 ± 16 | 366 | 321 ± 13 | 277 ± 8 | 0,153 | 2033 | 0,153 | 0,033 | 0,033 901 | 17 | 334 ± 42 | 297 ± 41 | 82 | 336 ± 27 | 290 ± 17 | 0,960 | 0,869 |
| 31–50 лет | 55 | 350 ± 21 | 120 | 324 ± 16 | 280 ± 13 | 0.054 | 0,010 | ||||||||||||
| 51–70 лет | 55 | 307 ± 23 | 281 ± 22 | 122 | 283 ± 12 | 248 ± 11 | 0,316 | 0,104 y | 24 | 264 ± 38 | 236 ± 30 | 42 | 243 ± 21 | 214 ± 21 | 0,631 | 0,541 | |||
| Женщины | 901 901 | 901 | |||||||||||||||||
| Кавказцы | 657 | 273 ± 7 | 246 ± 6 | 325 | 226 ± 10 | 201 ± 10 | <0.001 | <0,001 | |||||||||||
| 20–30 лет | 117 | 276 ± 16 | 246 ± 14 | 86 | 211 ± 16 | 184 ± 17 | 0,004 | 0,007 –50 лет | 182 | 289 ± 15 | 260 ± 13 | 112 | 233 ± 10 | 209 ± 11 | 0,002 | 0,001 | |||
| 51–70 лет | 261 | 193 7 | 245 ± 6 | 73 | 243 ± 16 | 222 ± 15 | 0.169 | 0,186 | |||||||||||
| ≥71 лет | 165 | 239 ± 10 | 215 ± 8 | 54 | 203 ± 20 | 181 ± 15 | 0,082 | 167 | 213 ± 11 | 184 ± 9 | 272 | 194 ± 7 | 166 ± 7 | 0,141 | 0,096 | ||||
| 20–30 лет | 24 | 32 | 176 ± 41 | 49 | 204 ± 22 | 180 ± 18 | 0.716 | 0,939 | |||||||||||
| 31–50 лет | 71 | 214 ± 19 | 186 ± 14 | 111 | 204 ± 10 | 179 ± 9 | 0,648 | 0,661 | 1 70 лет55 | 217 ± 16 | 192 ± 13 | 68 | 169 ± 8 | 150 ± 8 | 0,009 | 0,009 | |||
| ≥71 лет | 17 | 160 ± 14 | 44 | 158 ± 11 | 102 ± 28 | 0.615 | 0,084 | ||||||||||||
| Американцы мексиканского происхождения | 278 | 249 ± 11 | 216 ± 12 | 351 | 237 ± 8 | 211 ± 7 | 0,256 | 2021 900–0,621 | 59 | 289 ± 22 | 255 ± 22 | 101 | 250 ± 15 | 227 ± 14 | 0,073 | 0,164 | |||
| 31–50 лет | 77 | 24132 | 77 | 24 216 ± 17 | 123 | 239 ± 16 | 210 ± 13 | 0.785 | 0,724 | ||||||||||
| 51–70 лет | 108 | 227 ± 23 | 192 ± 28 | 93 | 200 ± 11 | 176 ± 12 | 0,303 | 0,578 | 1 y* 34 | 193 ± 16 | 171 ± 13 | 34 | 218 ± 24 | 184 ± 25 | 0,364 | 0,539 | |||
| . | Пользователи . | Непользователи . | P означает . | P геометрические средства . | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | . | . | |||||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | . | Среднее значение 1 . | Среднее значение 2 . | . | . | |||||||||||
| мг / день | мг / день | ||||||||||||||||||
| Мужчины | 9032 901 | 901 901 | |||||||||||||||||
| Кавказцы | 517 | 368 ± 9 | 331 ± 7 | 412 | 333 ± 9 | 294 ± 9 | 0.015 | 0,001 | |||||||||||
| 20–30 лет | 52 | 320 ± 24 | 287 ± 22 | 85 | 335 ± 19 | 291 ± 21 | 0,625 | 0,905 | 50 лет132 | 391 ± 20 | 350 ± 12 | 135 | 362 ± 15 | 324 ± 14 | 0,229 | 0,160 | |||
| 51–70 лет | 193 36832 | 339 ± 11 | 111 | 297 ± 12 | 272 ± 12 | <0.001 | <0,001 | ||||||||||||
| ≥71 лет | 140 | 342 ± 19 | 302 ± 14 | 81 | 263 ± 19 | 213 ± 30 | 0,003 | 0,015 | 116 | 333 ± 19 | 284 ± 14 | 255 | 255 ± 13 | 199 ± 20 | 0,002 | 0,004 | |||
| 20–30 лет | 16 | 34 279 ± 55 | 43 | 291 ± 27 | 254 ± 22 | 0.462 | 0,659 | ||||||||||||
| 31–50 лет | 47 | 366 ± 30 | 317 ± 24 | 106 | 248 ± 19 | 185 ± 35 | 0,005 | 0,016 | 70 лет37 | 279 ± 21 | 255 ± 17 | 70 | 226 ± 9 | 175 ± 16 | 0,025 | 0,001 | |||
| ≥71 лет | 16 | 172 ± 30 | 36 | 236 ± 30 | 193 ± 24 | 0.379 | 0,600 | ||||||||||||
| Американцы мексиканского происхождения | 151 | 355 ± 16 | 320 ± 16 | 366 | 321 ± 13 | 277 ± 8 | 0,153 | 2033 | 0,153 | 0,033 | 0,033 901 | 17 | 334 ± 42 | 297 ± 41 | 82 | 336 ± 27 | 290 ± 17 | 0,960 | 0,869 |
| 31–50 лет | 55 | 350 ± 21 | 120 | 324 ± 16 | 280 ± 13 | 0.054 | 0,010 | ||||||||||||
| 51–70 лет | 55 | 307 ± 23 | 281 ± 22 | 122 | 283 ± 12 | 248 ± 11 | 0,316 | 0,104 y | 24 | 264 ± 38 | 236 ± 30 | 42 | 243 ± 21 | 214 ± 21 | 0,631 | 0,541 | |||
| Женщины | 901 901 | 901 | |||||||||||||||||
| Кавказцы | 657 | 273 ± 7 | 246 ± 6 | 325 | 226 ± 10 | 201 ± 10 | <0.001 | <0,001 | |||||||||||
| 20–30 лет | 117 | 276 ± 16 | 246 ± 14 | 86 | 211 ± 16 | 184 ± 17 | 0,004 | 0,007 –50 лет | 182 | 289 ± 15 | 260 ± 13 | 112 | 233 ± 10 | 209 ± 11 | 0,002 | 0,001 | |||
| 51–70 лет | 261 | 193 7 | 245 ± 6 | 73 | 243 ± 16 | 222 ± 15 | 0.169 | 0,186 | |||||||||||
| ≥71 лет | 165 | 239 ± 10 | 215 ± 8 | 54 | 203 ± 20 | 181 ± 15 | 0,082 | 167 | 213 ± 11 | 184 ± 9 | 272 | 194 ± 7 | 166 ± 7 | 0,141 | 0,096 | ||||
| 20–30 лет | 24 | 32 | 176 ± 41 | 49 | 204 ± 22 | 180 ± 18 | 0.716 | 0,939 | |||||||||||
| 31–50 лет | 71 | 214 ± 19 | 186 ± 14 | 111 | 204 ± 10 | 179 ± 9 | 0,648 | 0,661 | 1 70 лет55 | 217 ± 16 | 192 ± 13 | 68 | 169 ± 8 | 150 ± 8 | 0,009 | 0,009 | |||
| ≥71 лет | 17 | 160 ± 14 | 44 | 158 ± 11 | 102 ± 28 | 0.615 | 0,084 | ||||||||||||
| Американцы мексиканского происхождения | 278 | 249 ± 11 | 216 ± 12 | 351 | 237 ± 8 | 211 ± 7 | 0,256 | 2021 900–0,621 | 59 | 289 ± 22 | 255 ± 22 | 101 | 250 ± 15 | 227 ± 14 | 0,073 | 0,164 | |||
| 31–50 лет | 77 | 24132 | 77 | 24 216 ± 17 | 123 | 239 ± 16 | 210 ± 13 | 0.785 | 0,724 | ||||||||||
| 51–70 лет | 108 | 227 ± 23 | 192 ± 28 | 93 | 200 ± 11 | 176 ± 12 | 0,303 | 0,578 | 1 y* 34 | 193 ± 16 | 171 ± 13 | 34 | 218 ± 24 | 184 ± 25 | 0,364 | 0,539 | |||
Не скорректированная диета , статус пользователя минералов или пищевых добавок, Национальное обследование здоровья и питания, 1999–2000 гг.
| . | Пользователи . | Непользователи . | P означает . | P геометрические средства . | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | . | . | |||||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | . | Среднее значение 1 . | Среднее значение 2 . | . | . | |||||||||||
| мг / день | мг / день | ||||||||||||||||||
| Мужчины | 9032 901 | 901 901 | |||||||||||||||||
| Кавказцы | 517 | 368 ± 9 | 331 ± 7 | 412 | 333 ± 9 | 294 ± 9 | 0.015 | 0,001 | |||||||||||
| 20–30 лет | 52 | 320 ± 24 | 287 ± 22 | 85 | 335 ± 19 | 291 ± 21 | 0,625 | 0,905 | 50 лет132 | 391 ± 20 | 350 ± 12 | 135 | 362 ± 15 | 324 ± 14 | 0,229 | 0,160 | |||
| 51–70 лет | 193 36832 | 339 ± 11 | 111 | 297 ± 12 | 272 ± 12 | <0.001 | <0,001 | ||||||||||||
| ≥71 лет | 140 | 342 ± 19 | 302 ± 14 | 81 | 263 ± 19 | 213 ± 30 | 0,003 | 0,015 | 116 | 333 ± 19 | 284 ± 14 | 255 | 255 ± 13 | 199 ± 20 | 0,002 | 0,004 | |||
| 20–30 лет | 16 | 34 279 ± 55 | 43 | 291 ± 27 | 254 ± 22 | 0.462 | 0,659 | ||||||||||||
| 31–50 лет | 47 | 366 ± 30 | 317 ± 24 | 106 | 248 ± 19 | 185 ± 35 | 0,005 | 0,016 | 70 лет37 | 279 ± 21 | 255 ± 17 | 70 | 226 ± 9 | 175 ± 16 | 0,025 | 0,001 | |||
| ≥71 лет | 16 | 172 ± 30 | 36 | 236 ± 30 | 193 ± 24 | 0.379 | 0,600 | ||||||||||||
| Американцы мексиканского происхождения | 151 | 355 ± 16 | 320 ± 16 | 366 | 321 ± 13 | 277 ± 8 | 0,153 | 2033 | 0,153 | 0,033 | 0,033 901 | 17 | 334 ± 42 | 297 ± 41 | 82 | 336 ± 27 | 290 ± 17 | 0,960 | 0,869 |
| 31–50 лет | 55 | 350 ± 21 | 120 | 324 ± 16 | 280 ± 13 | 0.054 | 0,010 | ||||||||||||
| 51–70 лет | 55 | 307 ± 23 | 281 ± 22 | 122 | 283 ± 12 | 248 ± 11 | 0,316 | 0,104 y | 24 | 264 ± 38 | 236 ± 30 | 42 | 243 ± 21 | 214 ± 21 | 0,631 | 0,541 | |||
| Женщины | 901 901 | 901 | |||||||||||||||||
| Кавказцы | 657 | 273 ± 7 | 246 ± 6 | 325 | 226 ± 10 | 201 ± 10 | <0.001 | <0,001 | |||||||||||
| 20–30 лет | 117 | 276 ± 16 | 246 ± 14 | 86 | 211 ± 16 | 184 ± 17 | 0,004 | 0,007 –50 лет | 182 | 289 ± 15 | 260 ± 13 | 112 | 233 ± 10 | 209 ± 11 | 0,002 | 0,001 | |||
| 51–70 лет | 261 | 193 7 | 245 ± 6 | 73 | 243 ± 16 | 222 ± 15 | 0.169 | 0,186 | |||||||||||
| ≥71 лет | 165 | 239 ± 10 | 215 ± 8 | 54 | 203 ± 20 | 181 ± 15 | 0,082 | 167 | 213 ± 11 | 184 ± 9 | 272 | 194 ± 7 | 166 ± 7 | 0,141 | 0,096 | ||||
| 20–30 лет | 24 | 32 | 176 ± 41 | 49 | 204 ± 22 | 180 ± 18 | 0.716 | 0,939 | |||||||||||
| 31–50 лет | 71 | 214 ± 19 | 186 ± 14 | 111 | 204 ± 10 | 179 ± 9 | 0,648 | 0,661 | 1 70 лет55 | 217 ± 16 | 192 ± 13 | 68 | 169 ± 8 | 150 ± 8 | 0,009 | 0,009 | |||
| ≥71 лет | 17 | 160 ± 14 | 44 | 158 ± 11 | 102 ± 28 | 0.615 | 0,084 | ||||||||||||
| Американцы мексиканского происхождения | 278 | 249 ± 11 | 216 ± 12 | 351 | 237 ± 8 | 211 ± 7 | 0,256 | 2021 900–0,621 | 59 | 289 ± 22 | 255 ± 22 | 101 | 250 ± 15 | 227 ± 14 | 0,073 | 0,164 | |||
| 31–50 лет | 77 | 24132 | 77 | 24 216 ± 17 | 123 | 239 ± 16 | 210 ± 13 | 0.785 | 0,724 | ||||||||||
| 51–70 лет | 108 | 227 ± 23 | 192 ± 28 | 93 | 200 ± 11 | 176 ± 12 | 0,303 | 0,578 | 1 y* 34 | 193 ± 16 | 171 ± 13 | 34 | 218 ± 24 | 184 ± 25 | 0,364 | 0,539 | |||
| . | Пользователи . | Непользователи . | P означает . | P геометрические средства . | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | n . | Потребление магния . | n . | Потребление магния . | . | . | |||||||||||||
| . | . | Среднее значение 1 . | Среднее значение 2 . | . | Среднее значение 1 . | Среднее значение 2 . | . | . | |||||||||||
| мг / день | мг / день | ||||||||||||||||||
| Мужчины | 9032 901 | 901 901 | |||||||||||||||||
| Кавказцы | 517 | 368 ± 9 | 331 ± 7 | 412 | 333 ± 9 | 294 ± 9 | 0.015 | 0,001 | |||||||||||
| 20–30 лет | 52 | 320 ± 24 | 287 ± 22 | 85 | 335 ± 19 | 291 ± 21 | 0,625 | 0,905 | 50 лет132 | 391 ± 20 | 350 ± 12 | 135 | 362 ± 15 | 324 ± 14 | 0,229 | 0,160 | |||
| 51–70 лет | 193 36832 | 339 ± 11 | 111 | 297 ± 12 | 272 ± 12 | <0.001 | <0,001 | ||||||||||||
| ≥71 лет | 140 | 342 ± 19 | 302 ± 14 | 81 | 263 ± 19 | 213 ± 30 | 0,003 | 0,015 | 116 | 333 ± 19 | 284 ± 14 | 255 | 255 ± 13 | 199 ± 20 | 0,002 | 0,004 | |||
| 20–30 лет | 16 | 34 279 ± 55 | 43 | 291 ± 27 | 254 ± 22 | 0.462 | 0,659 | ||||||||||||
| 31–50 лет | 47 | 366 ± 30 | 317 ± 24 | 106 | 248 ± 19 | 185 ± 35 | 0,005 | 0,016 | 70 лет37 | 279 ± 21 | 255 ± 17 | 70 | 226 ± 9 | 175 ± 16 | 0,025 | 0,001 | |||
| ≥71 лет | 16 | 172 ± 30 | 36 | 236 ± 30 | 193 ± 24 | 0.379 | 0,600 | ||||||||||||
| Американцы мексиканского происхождения | 151 | 355 ± 16 | 320 ± 16 | 366 | 321 ± 13 | 277 ± 8 | 0,153 | 2033 | 0,153 | 0,033 | 0,033 901 | 17 | 334 ± 42 | 297 ± 41 | 82 | 336 ± 27 | 290 ± 17 | 0,960 | 0,869 |
| 31–50 лет | 55 | 350 ± 21 | 120 | 324 ± 16 | 280 ± 13 | 0.054 | 0,010 | ||||||||||||
| 51–70 лет | 55 | 307 ± 23 | 281 ± 22 | 122 | 283 ± 12 | 248 ± 11 | 0,316 | 0,104 y | 24 | 264 ± 38 | 236 ± 30 | 42 | 243 ± 21 | 214 ± 21 | 0,631 | 0,541 | |||
| Женщины | 901 901 | 901 | |||||||||||||||||
| Кавказцы | 657 | 273 ± 7 | 246 ± 6 | 325 | 226 ± 10 | 201 ± 10 | <0.001 | <0,001 | |||||||||||
| 20–30 лет | 117 | 276 ± 16 | 246 ± 14 | 86 | 211 ± 16 | 184 ± 17 | 0,004 | 0,007 –50 лет | 182 | 289 ± 15 | 260 ± 13 | 112 | 233 ± 10 | 209 ± 11 | 0,002 | 0,001 | |||
| 51–70 лет | 261 | 193 7 | 245 ± 6 | 73 | 243 ± 16 | 222 ± 15 | 0.169 | 0,186 | |||||||||||
| ≥71 лет | 165 | 239 ± 10 | 215 ± 8 | 54 | 203 ± 20 | 181 ± 15 | 0,082 | 167 | 213 ± 11 | 184 ± 9 | 272 | 194 ± 7 | 166 ± 7 | 0,141 | 0,096 | ||||
| 20–30 лет | 24 | 32 | 176 ± 41 | 49 | 204 ± 22 | 180 ± 18 | 0.716 | 0,939 | |||||||||||
| 31–50 лет | 71 | 214 ± 19 | 186 ± 14 | 111 | 204 ± 10 | 179 ± 9 | 0,648 | 0,661 | 1 70 лет55 | 217 ± 16 | 192 ± 13 | 68 | 169 ± 8 | 150 ± 8 | 0,009 | 0,009 | |||
| ≥71 лет | 17 | 160 ± 14 | 44 | 158 ± 11 | 102 ± 28 | 0.615 | 0,084 | ||||||||||||
| Американцы мексиканского происхождения | 278 | 249 ± 11 | 216 ± 12 | 351 | 237 ± 8 | 211 ± 7 | 0,256 | 2021 900–0,621 | 59 | 289 ± 22 | 255 ± 22 | 101 | 250 ± 15 | 227 ± 14 | 0,073 | 0,164 | |||
| 31–50 лет | 77 | 24132 | 77 | 24 216 ± 17 | 123 | 239 ± 16 | 210 ± 13 | 0.785 | 0,724 | ||||||||||
| 51–70 лет | 108 | 227 ± 23 | 192 ± 28 | 93 | 200 ± 11 | 176 ± 12 | 0,303 | 0,578 | 1 y* 34 | 193 ± 16 | 171 ± 13 | 34 | 218 ± 24 | 184 ± 25 | 0,364 | 0,539 | |||
Мы также исследовали источники питания на предмет потребления магния.Участники сообщили о 3719 различных продуктах, содержащих диетический магний. Согласно пищевым кодексам, 10 основных вкладчиков в потребление магния с пищей составляли молотый кофе (3,7% от потребления), 2% молоко (2,6%), цельное молоко (2,1%), сырые бананы (1,8%), пиво (1,8%). ), обезжиренное или обезжиренное молоко (1,5%), апельсиновый сок (1,5%), легкое пиво (1,4%), картофель фри (1,1%) и 1% молоко (0,9%). Когда мы сгруппировали пищевые коды по широким категориям, в первую десятку вошли овощи (12,9%), молоко (7,5%), мясо (7,3%), хлеб (6.6%), смешанные блюда (6,5%), пицца, паста и другие разные блюда (5,6%), кофе (5,3%), сладости (4,6%), холодные каши (4,1%) и алкогольные напитки (3,8%). Смешанные блюда включали блюда, приготовленные из мяса или рыбы и овощей. Кроме того, на комбинированную категорию фруктов и фруктовых соков приходилось 6,3% потребления магния.
1″> ЦИТИРОВАННАЯ ЛИТЕРАТУРА
1.Fox
,C.
,Ramsoomair
,D.
иCarter
,C.
(2001
)Магний: его доказанное и потенциальное клиническое значение
.Юг. Med. J.
94
:1195
–1201
.2.Капуччио
,F. P.
(2000
)Натрий, калий, кальций и магний и риск сердечно-сосудистых заболеваний
.J. Cardiovasc. Риск
7
:1
–3
.3.Mizushima
,S.
,Cappuccio
,F. P.
,Nichols
,R.
иElliott
,P.
(1998
)Диетическое потребление магния и артериальное давление: качественный обзор обсервационных исследований
.J. Hum. Гипертензии.
12
:447
–453
.4.Kao
,WH
,Folsom
,AR
,Nieto
,FJ
,Mo
,JP
,Watson
,RL
и1999
)Сыворотка и диетический магний и риск сахарного диабета 2 типа: исследование риска атеросклероза в сообществах
.Arch. Междунар. Med.
159
:2151
–2159
. 5.Altura
,B. M.
&Altura
,B. T.
(2001
)Головные боли напряжения и напряжение мышц: есть ли роль магния?
.Med. Гипотезы
57
:705
–713
.6.Weaver
,C. M.
(2000
)(2000) Потребность детей и подростков в кальции и магнии и максимальная костная масса
.Питание
16
:514
–516
.7.Лукаски
,Х.С.
(2001
)Магний, цинк и хром, питание и спортивные результаты
.Can J. Appl. Physiol.
26
(доп.):S13
–S22
.8.Ceremuzynski
,L.
,Gebalska
,J.
,Wolk
,R.
&Makowska
,E.
(2000
)Гипомагниемия при сердечной недостаточности с желудочковыми аритмиями. Благоприятные эффекты от приема магния
.J. Intern. Med.
247
:78
–86
.9.Shechter
,M.
,Sharir
,M.
,Labrador
,MJ
,Forrester
,J.
,Silver
,B.
& MerBairey
С.N.
(2000
)Пероральная терапия магнием улучшает эндотелиальную функцию у пациентов с ишемической болезнью сердца
.Тираж
102
:2353
–2358
. 10.Jee
,SH
,Miller
,ER
,3rd, Guallar
,E.
,Singh
,VK
,Appel
,LJ
& (2002
)Влияние добавок магния на артериальное давление: метаанализ рандомизированных клинических исследований
.г. J. Hypertens.
15
:691
–696
. 11.de Valk
,H. W.
(1999
)Магний при сахарном диабете
.Neth. J. Med.
54
:139
–146
.12.Hill
,J.
,Micklewright
,A.
,Lewis
,S.
иBritton
,J.
(1997
)Расследование последствий короткого изменение срока потребления магния с пищей при астме
.евро. Респир. J.
10
:2225
–2229
. 13.Маускоп
,A.
иAltura
,B. M.
(1998
)Роль магния в патогенезе и лечении мигрени
.Clin. Neurosci.
5
:24
–27
. 14.Makrides
,M.
иCrowther
,C.A.
(2001
)Добавки магния во время беременности
.Кокрановская база данных Syst. Ред.
4
CD000937.15.Swain
,R.
иKaplan-Machlis
,B.
(1999
)Магний для следующего тысячелетия
.Юг. Med. J.
92
:1040
–1047
. 16.Dacey
,M.J.
(2001
)Гипомагниемия
.Крит. Care Clin.
17
:155
–173
.17.Pennington
,J. A.
&Schoen
,S. A.
(1996
)Общее исследование диеты: оценочное потребление питательных элементов с пищей, 1982–1991
.Внутр. J. Vitam. Nutr. Res.
66
:350
–362
. 18.Galan
,P.
,Preziosi
,P.
,Durlach
,V.
,Valeix
,P.
,Ribas
,L.
,Bouzid
,D.
,Favier
,A.
иHercberg
,S.
(1997
)Потребление магния с пищей взрослым французским населением
.Магн. Res.
10
:321
–328
.19.Wilson
,J. W.
,Enns
,C. W.
,Goldman
,J. D.
,Tippett
,K. S.
,Mickle
,S.J.
,Cleveland
,LE
иChahil
,PS
(1997
)Таблицы данных: объединенные результаты непрерывного исследования потребления пищевых продуктов отдельными лицами, проведенного Министерством сельского хозяйства США в 1994 и 1995 годах, и исследования знаний в области здравоохранения в 1994 и 1995 годах.
.Министерство сельского хозяйства США, Служба сельскохозяйственных исследований
1997
Исследовательская группа продовольственных исследований
Белтсвилл, Мэриленд
. http://www.barc.usda.gov/bhnrc/foodsurvey/pdf/Tbchts95.pdf (последний просмотр 3 марта 2003 г.) 20.Ford
,E. S.
(1998
)Раса, образование и диетические катионы: результаты Третьего национального исследования здоровья и питания
.Этн. Дис.
8
:10
–20
. 21.Vaquero
,M. P.
(2002
)Магний и микроэлементы у пожилых людей: потребление, статус и рекомендации
.J. Nutr. Здоровье Старение
6
:147
–153
.22.Iannello
,S.
&Belfiore
,F.
(2001
)Гипомагниемия. Обзор патофизиологических, клинических и терапевтических аспектов
.Panminerva Med.
43
:177
–209
. 23.Lowenstein
,F. W.
иStanton
,M. F.
(1986
)Уровни магния в сыворотке крови в США, 1971–1974
.J. Am. Coll. Nutr.
5
:399
–414
. 24.Ford
,E. S.
(1999
)Сывороточный магний и ишемическая болезнь сердца: результаты национальной выборки среди взрослых в США
.Внутр. J. Epidemiol.
28
:645
–651
. 25.Whang
,R.
иSims
,G.
(2000
)Добавки магния и калия для профилактики диабетических сосудистых заболеваний
.Med. Гипотезы
55
:263
–265
. 29.Институт медицины
(1997
)Нормы потребления кальция, фосфора, магния, витамина D и фторида с пищей
1997
National Academy Press
Вашингтон, округ Колумбия
.30.Innerarity
,S.
(2000
)Гипомагниемия при острых и хронических заболеваниях
.Крит. Care Nurs.В.
23
:1
–19
. 31.Zemel
,P.
,Gualdoni
,S.
&Sowers
,J. R.
(1988
)Расовые различия в потреблении минералов при амбулаторном нормотензивном и гипертензивном
.г. J. Hypertens.
1
:146S
–148S
.32.Гербер
,А. М.
,Джеймс
,С. А.
,Аммерман
,А.S.
,Keenan
,NL
,Garrett
,JM
,Strogatz
,DS
иHaines
,PS
(1991
teconomic) ПотреблениеСоциально-экономичный : Исследование округа Питт
.г. J. Общественное здравоохранение
81
:1608
–1612
. 33.Саймон
,J. A.
,Obarzanek
,E.
,Daniels
,S.R.
иFrederick
,M. M.
(1994
)Потребление катионов в пище и артериальное давление у чернокожих и белых девочек
.г. J. Epidemiol.
139
:130
–140
. 34.Министерство сельского хозяйства США
(1995
)Потребление пищи и питательных веществ отдельными лицами в Соединенных Штатах, 1 день, 1989–91. Продолжающееся обследование потребления пищи отдельными лицами, 1989–91. Отчет о национальных исследованиях в области питания №91–92
1995
Министерство сельского хозяйства США, Служба сельскохозяйственных исследований
Riverdale, MD
.35.Ma
,J.
,Folsom
,AR
,Melnick
,SL
,Eckfeldt
,JH
,Sharrett
,AR
,Hutchinson
,RG
иMetcalf
,P.A.
(1995
)Связь сывороточного и диетического магния с сердечно-сосудистыми заболеваниями, гипертонией, диабетом, инсулином и толщиной стенки сонной артерии: исследование ARIC
.J. Clin. Эпидемиол.
48
:927
–940
,36.Lopez
,T. K.
,Marshall
,J. A.
,Shetterly
,S. M.
,Baxter
,J.
иHamman
,R.F.
(1995
)Этнические различия в потреблении микронутриентов среди сельского двухэтнического населения
.г. J. Prev. Med.
11
:301
–305
0,37.Исследовательская группа по исследованиям пищевых продуктов Министерства сельского хозяйства США
(1998
)Таблицы данных: потребление пищевых продуктов и питательных веществ по расам
1998
:1994
–1996
Служба сельскохозяйственных исследований Министерства сельского хозяйства США
,Белтсвилл, Мэриленд
.Интернет: http://www.barc.usda.gov/bhnrc/foodsurvey/pdf/Race.pdf (по состоянию на 3 марта 2003 г.) 38.Thompson
,F. E.
иByers
,T.
(1994
)Руководство по оценке питания
.г. J. Clin. Nutr.
124
(доп.):2245S
–2317S
.39.Валлийский
,S.
,Дэвис
,C.
иShaw
,A.
(1992
)Разработка пирамиды Food Guide
.Nutr. Сегодня
27
:12
–18
.Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Вариант h4.3 гистона h4 регулирует метилирование ДНК тела гена у Arabidopsis thaliana | Геномная биология
Talbert PB, Henikoff S. Варианты гистонов в движении: субстраты для динамики хроматина. Nat Rev Mol Cell Biol. 2017; 18: 115–26.
CAS Статья PubMed Google ученый
Филипеску Д., Мюллер С., Альмоузни Г. Варианты гистона h4 и их шапероны во время развития и болезни: вклад в эпигенетический контроль. Annu Rev Cell Dev Biol. 2014; 30: 615–46.
CAS Статья PubMed Google ученый
Talbert PB, Ahmad K, Almouzni G, Ausió J, Berger F, Bhalla PL и др. Единая номенклатура вариантов гистонов, основанная на филогенезе. Эпигенетика хроматина. 2012; 5: 7.
CAS Статья PubMed Central PubMed Google ученый
Ахмад К., Хеникофф С. Вариант гистона h4.3 маркирует активный хроматин путем независимой от репликации сборки нуклеосом. Mol Cell. 2002; 9: 1191–200.
CAS Статья PubMed Google ученый
Hodl M, Basler K. Транскрипция в отсутствие гистона h4.3. Curr Biol. 2009; 19: 1221–6.
CAS Статья PubMed Google ученый
Сакаи А., Шварц Б.Е., Гольдштейн С., Ахмад К. Транскрипционные и онтогенетические функции варианта гистона h4.3 у Drosophila . Curr Biol. 2009; 19: 1816–20.
CAS Статья PubMed Central PubMed Google ученый
Couldrey C, Carlton MB, Nolan PM, Colledge WH, Evans MJ. Вставка ловушки ретровирусного гена в ген гистона 3.3A вызывает частичную неонатальную летальность, задержку роста, нервно-мышечный дефицит и самцовскую субфертильность у трансгенных мышей. Hum Mol Genet. 1999; 8: 2489–95.
CAS Статья PubMed Google ученый
Szenker E, Lacoste N, Almouzni G. Потребность в развитии HIRA-зависимого отложения h4.3, выявленная при гаструляции у Xenopus .Cell Rep. 2012; 1: 730–40.
CAS Статья PubMed Google ученый
Акияма Т., Сузуки О., Мацуда Дж., Аоки Ф. Динамическая замена вариантов гистона h4 репрограммирует эпигенетические метки в ранних эмбрионах мыши. PLoS Genet. 2011; 7: e1002279.
CAS Статья PubMed Central PubMed Google ученый
Джуллиен Дж., Астранд С., Зенкер Э., Гарретт Н., Альмоузни Дж., Гурдон Дж. Б..HIRA-зависимое отложение h4.3 необходимо для транскрипционного репрограммирования после переноса ядра в ооциты Xenopus . Эпигенетика хроматина. 2012; 5: 17.
CAS Статья PubMed Central PubMed Google ученый
Loppin B, Bonnefoy E, Anselme C, Laurencon A, Karr TL, Couble P. Шаперон гистона h4.3 HIRA необходим для сборки хроматина в мужском пронуклеусе. Природа. 2005; 437: 1386–90.
CAS Статья PubMed Google ученый
Орси Г.А., Альгазери А., Мейер Р.Э., Капри М., Сапей-Триумф Л.М., Хорард Б. и др. Йемануклеин дрозофилы и HIRA кооперируются для сборки de novo h4.3-содержащих нуклеосом в мужском пронуклеусе. PLoS Genet. 2013; 9: e1003285.
CAS Статья PubMed Central PubMed Google ученый
Santenard A, Torres-Padilla M-E.Эпигенетическое репрограммирование в репродукции млекопитающих: вклад вариантов гистонов. Эпигенетика. 2009; 4: 80–4.
CAS Статья PubMed Google ученый
Ингуфф М., Радемахер С., Холек С., Шолич Л., Синь Н., Ридшоу А. и др. Зиготическая перезагрузка репертуара вариантов HISTONE 3 участвует в эпигенетическом репрограммировании у арабидопсиса . Curr Biol. 2010; 20: 2137–43.
CAS Статья PubMed Google ученый
Nie X, Wang H, Li J, Holec S, Berger F. Комплекс HIRA, который откладывает гистон h4.3, консервативен в Arabidopsis и способствует динамике транскрипции. Биол Открытый. 2014; 3: 794–802.
CAS Статья PubMed Central PubMed Google ученый
McKittrick E, Gafken PR, Ahmad K, Henikoff S. Гистон h4.3 обогащен ковалентными модификациями, связанными с активным хроматином. Proc Natl Acad Sci U S A.2004; 101: 1525–30.
CAS Статья PubMed Central PubMed Google ученый
Шу Х., Накамура М., Сирецкий А., Борги Л., Мораес И., Вильдхабер Т. и др. Arabidopsis вариант замены гистона h4.3 занимает промоторы регулируемых генов. Genome Biol. 2014; 15: R62.
Артикул PubMed Central PubMed Google ученый
Страуд Н., Отеро С., Десвойес Б., Рамирес-Парра Е., Якобсен С.Е., Гутьеррес С. Полногеномный анализ вариантов гистона h4.1 и h4.3 в Arabidopsis thaliana . Proc Natl Acad Sci U S. A. 2012; 109: 5370–5.
CAS Статья PubMed Central PubMed Google ученый
Wollmann H, Holec S, Alden K, Clarke ND, Jacques PE, Berger F. Динамическое отложение варианта гистона h4.3 сопровождает онтогенетическое ремоделирование транскриптома Arabidopsis .PLoS Genet. 2012; 8: e1002658.
CAS Статья PubMed Central PubMed Google ученый
Голдберг А.Д., Банашинский Л.А., Но К.М., Льюис П.В., Эльзэссер С.Дж., Штадлер С. и др. Различные факторы контролируют вариант гистона h4. 3 локализация в определенных регионах генома. Клетка. 2010; 140: 678–91.
CAS Статья PubMed Central PubMed Google ученый
Ray-Gallet D, Woolfe A, Vassias I, Pellentz C, Lacoste N, Puri A, et al. Динамика отложения гистона h4 in vivo выявляет механизм заполнения нуклеосомных промежутков для h4.3, чтобы поддерживать целостность хроматина. Mol Cell. 2011; 44: 928–41.
CAS Статья PubMed Google ученый
Banaszynski LA, Wen D, Dewell S, Whitcomb SJ, Lin M, Diaz N, et al. Hira-зависимое отложение гистона h4.3 облегчает рекрутирование prc2 в онтогенетические локусы в ES-клетках.Клетка. 2013; 155: 107–20.
CAS Статья PubMed Google ученый
Земах А., МакДэниэл И.Е., Сильва П., Зильберман Д. Полногеномный эволюционный анализ метилирования эукариотической ДНК. Наука. 2010; 328: 916–9.
CAS Статья PubMed Google ученый
Зильберман Д., Геринг М., Тран Р.К., Баллинджер Т., Хеникофф С. Полногеномный анализ метилирования ДНК Arabidopsis thaliana обнаруживает взаимозависимость между метилированием и транскрипцией.Нат Жене. 2007; 39: 61–9.
CAS Статья PubMed Google ученый
To TK, Saze H, Kakutani T. Метилирование ДНК в транскрибируемых областях. Plant Physiol. 2015; 168: 1219–25.
CAS Статья PubMed Central PubMed Google ученый
Баубек Т., Коломбо Д.Ф., Вирбелауэр С., Шмидт Дж., Бургер Л., Кребс А.Р. и др. Геномное профилирование ДНК-метилтрансфераз показывает роль DNMT3B в метилировании генов.Природа. 2015; 520: 243–7.
CAS Статья PubMed Google ученый
Вагнер Э.Дж., Карпентер ПБ. Понимание языка метилирования Lys36 гистона h4. Nat Rev Mol Cell Biol. 2012; 13: 115–26.
CAS Статья PubMed Central PubMed Google ученый
Бьюик А.Дж., Джи Л., Нидерхут С.Е., Уиллинг Э.М., Хофмайстер Б.Т., Ши Х и др.О происхождении и эволюционных последствиях метилирования ДНК тела гена. Proc Natl Acad Sci U S. A. 2016; 113: 9111–6.
CAS Статья PubMed Central PubMed Google ученый
Такуно С., Ран Дж. Х., Гаут Б.С. Эволюционные паттерны метилирования генной ДНК различаются в зависимости от наземных растений. Nat Plants. 2016; 2: 15222.
CAS Статья PubMed Google ученый
Видалис А., Живкович Д., Варденаар Р., Рокис Д., Телье А., Йоханнес Ф. Эволюция метиломов у растений. Genome Biol. 2016; 17: 264.
Артикул PubMed Central PubMed Google ученый
Ausin I., Feng S, Yu C., Liu W, Kuo HY, Jacobsen EL, et al. Метилом ДНК 20-гигабазного генома ели европейской. Proc Natl Acad Sci U S. A. 2016; 113: E8106–13.
CAS Статья PubMed Central PubMed Google ученый
Okada T, Endo M, Singh MB, Bhalla PL. Анализ семейства генов гистона h4 в Arabidopsis и идентификация специфичного для мужских гамет варианта AtMGh4. Плант Дж. 2005; 44: 557–68.
CAS Статья PubMed Google ученый
Рудье Ф., Ахмед И., Берар С., Саразин А., Мэри-Юард Т., Кортихо С. и др. Интегративное эпигеномное картирование определяет четыре основных состояния хроматина в Arabidopsis . EMBO J.2011; 30: 1928–38.
CAS Статья PubMed Central PubMed Google ученый
Асейтуно Ф.Ф., Мосейко Н., Ри С.Ю., Гутьеррес РА. Правила экспрессии генов в растениях: идентичность органов и метилирование тел генов являются ключевыми факторами регуляции экспрессии генов у Arabidopsis thaliana . BMC Genomics. 2008; 9: 438.
Артикул PubMed Central PubMed Google ученый
Cokus SJ, Feng S, Zhang X, Chen Z, Merriman B, Haudenschild CD и др. Бисульфитное секвенирование генома Arabidopsis выявляет паттерн метилирования ДНК. Природа. 2008; 452: 215–9.
CAS Статья PubMed Central PubMed Google ученый
Lister R, O’Malley RC, Tonti-Filippini J, Gregory BD, Berry CC, Millar AH, et al. Высокоинтегрированные карты эпигенома с одним базовым разрешением Arabidopsis .Клетка. 2008. 133: 523–36.
CAS Статья PubMed Central PubMed Google ученый
Чжан Х, Язаки Дж., Сундаресан А., Кокус С., Чан С.В., Чен Х. и др. Полногеномное картирование с высоким разрешением и функциональный анализ метилирования ДНК в Arabidopsis . Клетка. 2006; 126: 1189–201.
CAS Статья PubMed Google ученый
Тран Р.К., Хеникофф Дж. Г., Зильберман Д., Дитт Р.Ф., Якобсен С.Е., Хеникофф С.Профилирование метилирования ДНК позволяет идентифицировать кластеры метилирования CG в генах Arabidopsis . Curr Biol. 2005; 15: 154–9.
CAS Статья PubMed Google ученый
Зильберман Д., Коулман-Дерр Д., Баллинджер Т., Хеникофф С. Гистон h3A.Z и метилирование ДНК являются взаимно антагонистическими метками хроматина. Природа. 2008; 456: 125–9.
CAS Статья PubMed Central PubMed Google ученый
Коулман-Дерр Д., Зильберман Д. Отложение гистона варината h3A.Z в телах генов регулирует гены ответа. PLoS Genet. 2012; 8 (10): e1002988.
CAS Статья PubMed Central PubMed Google ученый
Woodcock CL, Skoultchi AI, Fan Y. Роль линкерного гистона в структуре и функции хроматина: стехиометрия h2 и длина нуклеосомного повтора. Chromosom Res. 2006; 14: 17–25.
CAS Статья Google ученый
Хергет С.П., Шнайдер Р. Линкерные гистоны h2: многофункциональные белки за пределами ядерной ядерной частицы. EMBO Rep. 2015; 16: 1439–53.
CAS Статья PubMed Central PubMed Google ученый
Routh A, Sandin S, Rhodes D. Длина нуклеосомного повтора и стехиометрия линкерного гистона определяют структуру волокна хроматина. Proc Natl Acad Sci U S. A. 2008; 105: 8872–7.
CAS Статья PubMed Central PubMed Google ученый
Земах А., Ким М.Ю., Се П.Х., Коулман-Дерр Д., Эшед-Вильямс Л., Тао К. и др. Ремоделирующий нуклеосомы Arabidopsis DDM1 позволяет ДНК-метилтрансферазам получать доступ к h2-содержащему гетерохроматину. Клетка. 2013; 153: 193–205.
CAS Статья PubMed Central PubMed Google ученый
Jin C, Felsenfeld G. Стабильность нуклеосом, опосредованная вариантами гистонов h4.3 и h3A.Z. Genes Dev. 2007; 21: 1519–29.
CAS Статья PubMed Central PubMed Google ученый
Коулман-Дерр Д., Зильберман Д. Метилирование ДНК, h3A.Z и регуляция конститутивной экспрессии. Колд Спринг Харб Symp Quant Biol. 2012; 77: 147–54.
CAS Статья PubMed Google ученый
Страуд Х., Гринберг М.В.К., Фенг С., Бернатавичуте Ю.В., Якобсен С.Е. Всесторонний анализ сайленсирующих мутантов показывает сложную регуляцию метилома Arabidopsis .Клетка. 2013; 152: 352–64.
CAS Статья PubMed Central PubMed Google ученый
Braunschweig U, Hogan GJ, Pagie L, van Steensel B. Связывание гистона h2 ингибируется вариантом гистона h4.3. EMBO J. 2009; 28: 3635–45.
CAS Статья PubMed Central PubMed Google ученый
Lin C-J, Conti M, Ramalho-Santos M. Вариант гистона h4.3 поддерживает состояние деконденсированного хроматина, необходимое для доимплантационного развития мышей. Разработка. 2013; 140: 3624–34.
CAS Статья PubMed Central PubMed Google ученый
Робинсон П.Дж., Родс Д. Структура «30 нм» хроматинового волокна: ключевая роль линкерного гистона. Curr Opin Struct Biol. 2006; 16: 336–43.
CAS Статья PubMed Google ученый
Henikoff S. Лабильные нуклеосомы h4.3 + h3A.Z маркируют «участки, свободные от нуклеосом». Нат Жене. 2009. 41: 865–6.
CAS Статья PubMed Google ученый
Tsutsui H, Higashiyama T. Векторы pKAMA-ITACHI для высокоэффективного CRISPR / Cas9-опосредованного нокаута гена в Arabidopsis thaliana . Physiol растительной клетки. 2017; 58: 46–56.
PubMed Google ученый
Schwab R, Ossowski S, Riester M, Warthmann N, Weigel D. Высокоспецифичное подавление генов с помощью искусственных микроРНК в Arabidopsis . Растительная клетка. 2006; 5: 1121–33.
Артикул Google ученый
Yelagandula R, Stroud H, Holec S, Zhou K, Feng S, Zhong X и др. Вариант гистона h3A.W определяет гетерохроматин и способствует конденсации хроматина в Arabidopsis . Клетка. 2014; 158: 98–109.
CAS Статья PubMed Central PubMed Google ученый
da Huang W, Sherman BT, Lempicki RA. Инструменты обогащения биоинформатики: пути к всестороннему функциональному анализу больших списков генов. Nucleic Acids Res. 2009; 37: 1–13.
Артикул Google ученый
da Huang W, Sherman BT, Lempicki RA. Систематический и комплексный анализ больших списков генов с использованием ресурсов биоинформатики DAVID. Nat Protoc. 2009. 4: 44–57.
CAS Статья Google ученый
Wu Z, Irizarry RA. Предварительная обработка данных массива олигонуклеотидов. Nat Biotechnol. 2004. 22: 656–8.
CAS Статья PubMed Google ученый
Langmead B, Schatz MC, Lin J, Pop M, Salzberg SL. Поиск SNP с помощью облачных вычислений. Genome Biol. 2009; 10: R134.
Артикул PubMed Central PubMed Google ученый
Chen P-Y, Cokus SJ, Pellegrini M.BS Seeker: точное картирование для бисульфитного секвенирования. BMC Bioinformatics. 2010; 11: 203.
CAS Статья PubMed Central PubMed Google ученый
Подробный обзор выпуклых и вогнутых углов в объемной микрообработке кремния на основе анизотропного влажного химического травления | Письма о микро- и наносистемах
Гад-эль-Хак М. (2002) Справочник по МЭМС. CRC Press LLC, Бока-Ратон
MATH Google ученый
Элвенспук М., Янсен Х. (1998) Микрообработка кремния. Издательство Кембриджского университета, Великобритания
Google ученый
Такахата К. (2013) Достижения в области микро / нано-электромеханических систем и производственных технологий. Издатель: InTech.
Frühauf J (2005) Форма и функциональные элементы микротехники объемного кремния: руководство по кремниевым структурам с влажным травлением. Springer.
Линдроос В., Тилли М., Лехто А., Мотоока Т. (2010) Справочник по материалам и технологиям МЭМС на основе кремния.Издательство Уильям Эндрю.
Hsu TR (2003) МЭМС и микросистемы: проектирование и производство. Tata McGraw-Hill Publishing Company Ltd, Нью-Дели, Индия
Google ученый
Маду MJ (2002) Основы микротехнологии: Наука миниатюризации, 2-е изд. CRC Press, Бока-Ратон, США
Google ученый
Варадан В.К. (2006) Системы интеллектуальных материалов и МЭМС: методологии проектирования и разработки.John Wiley & Sons Ltd.
Bustillo JM, Howe RT, Muller RS (1998) Микрообработка поверхности для микроэлектромеханических систем. IEEE Proc 86: 1552–74
Google ученый
Ковач Г.Т., Малуф Н.И., Петерсен К.Е. (1998) Объемная микрообработка кремния. IEEE Proc 86: 1536–1351
Google ученый
Петерсен К.Э. (1982) Кремний как механический материал.IEEE Proc 70: 420–457
Google ученый
Ланг В. (1996) Технология микроструктурирования кремния. Материаловедение и инженерия: R: Отчеты 17: 1–55
Google ученый
Янсен Х., Гарденир Х., Бур М.Д., Элвенспук М., Флюитман Дж. (1996) Обзор реактивного ионного травления кремния в микротехнологии. J Micromech Microeng 6: 14–28
Google ученый
Oehrlein GS (1990) Реактивное ионное травление. В: Rossnagel SM, Westwood WD, Haber JJ (eds) Справочник по основам технологии плазменной обработки, травлению, осаждению и взаимодействию поверхностей. Нью-Джерси: Нойес, Парк Ридж
Google ученый
Coburn JW, Winters HF (1979) Плазменное травление – обсуждение механизмов. J Vac Sci Technol 16: 391–403
Google ученый
Лармер Ф., Шилп П. (1994) Метод анизотропного травления кремния, Патент Германии DE 4 241 045.
Цзян Э., Китинг А., Мартынюк М., Прасад К., Фараоне Л., Цзян Дж. М. (2012) низкотемпературная объемная микрообработка кремния с использованием индуктивно связанной плазмы SF 6 / O 2 . J Micromech Microeng 22: 0
(10pp)
Google ученый
Хайнс А.М., Ашраф Х., Бхардвадж Дж. К., Хопкинс Дж., Джонстон И., Шеперд Дж. Н. (1999) Последние достижения в травлении кремния для МЭМС с использованием процесса ASE.Актуаторы Sens A 74: 13–17
Google ученый
Teng J, Prewett PD (2005) Изготовление сфокусированным ионным пучком термически активируемых биморфных кантилеверов. Актуаторы Sens A 123–124: 608–613
Google ученый
Walker CK, Narayanan G, Knoepfle H, Capara J, Glenn J, Hungerford A, Bloomstein TM, Palmacci ST, Stern MB, Curtin JE (1997) Лазерная микрообработка кремния: новая техника для получения высокого качества компоненты терагерцового волновода.В: Proc. 8 -й Международный симпозиум по космическим терагерцовым технологиям. Гарвардский университет, Кембридж, США, 25-27 марта 1997 г., стр. 358. http://www.nrao.edu/meetings/isstt/papers/1997/19376.pdf
Schwartz B, Robbins H (1976) Химическое травление кремния. J Electrochem Soc 123 (12): 1903–1909
Google ученый
Занди К., Арзи Э., Изади Н., Мохаджерзаде С., Хаджи С., Абди Ю., Асл Сулеймани Э. (2006) Исследование объемной микрообработки кремния <100>.Eur Phys J Appl Phys 35: 7–12
Google ученый
Lee DB (1969) Анизотропное травление кремния. J Appl Phys 40: 4569–4575
Google ученый
Бин К.Е. (1978) Анизотропное травление кремния. IEEE Trans Electron Devices ED-25: 1185–1193
Google ученый
Зайдель Х., Чепреги Л., Хойбергер А., Баумгартель Х. (1990) Анизотропное травление кристаллического кремния в щелочных растворах I. Зависимость от ориентации и поведение пассивирующих слоев.J Electrochem Soc 137 (11): 3612–3626
Google ученый
Зайдель Х., Чепреги Л., Хойбергер А., Баумгартель Х. (1990) Анизотропное травление кристаллического кремния в щелочных растворах II: Влияние примесей. J Electrochem Soc 137: 3626–3632
Google ученый
Табата О., Асахи Р., Фунабаши Х, Шимаока К., Сугияма С. (1992) Анизотропное травление кремния в растворах ТМАГ.Актуаторы Sens A 34 (1): 51–57
Google ученый
Сато К., Шикида М., Мацусима Ю., Ямаширо Т., Асауми К., Ирие Ю., Ямамото М. (1998) Характеристика ориентационно-зависимых травильных свойств монокристаллического кремния: эффекты концентрации КОН. Актуаторы Sens A 61: 87–93
Google ученый
Сато К., Шикида М., Ямаширо Т., Цунекава М., Ито С. (1999) Придание шероховатости поверхности монокристаллического кремния, протравленной водным раствором КОН.Актуаторы Sens A 73: 122–130
Google ученый
Пауэлл О., Харрисон Х. Б. (2001) Анизотропное травление плоскостей {100} и {110} в кремнии (100). J Micromech Microeng 11: 217–220
Google ученый
Танака Х., Ямашита С., Абэ Й., Шикида М., Сато К. (2004) Быстрое травление кремния с гладкой поверхностью в высокотемпературных диапазонах вблизи точки кипения раствора КОН.Актуаторы Sens A 114: 516–520
Google ученый
Мацуока М., Йошида Ю., Моронуки М. (1992) Изготовление кремниевых тонких диафрагм, свободных от микропирамид, с использованием анизотропного травления в растворе КОН. J Chem Eng 25: 735–740
Google ученый
Барьека И., Зубель И. (1995) Анизотропное травление кремния в травителе КОН-изопропанол. Актуаторы Sens A 48: 229–238
Google ученый
Шикида М., Сато К., Токоро К., Учикава Д. (2000) Различия в свойствах анизотропного травления растворов КОН и ТМАГ. Актуаторы Sens A 80: 179–188
Google ученый
Backlund Y, Rosengren L (1992) Новые формы в (100) Si с использованием травления KOH и EDP. J Micromech Microeng 27: 5–9
Google ученый
Сато К., Шикида М., Ямаширо Т., Асауми К., Ирие Ю., Ямамото М. (1999) Скорость анизотропного травления монокристаллического кремния для водного раствора ТМАГ в зависимости от кристаллографической ориентации.Актуаторы Sens A 73: 131–137
Google ученый
Ченг Д., Госальвез М.А., Хори Т., Сато К., Шикида М. (2006) Улучшение гладкости анизотропно протравленных кремниевых поверхностей: влияние концентраций поверхностно-активного вещества и ТМАГ. Актуаторы Sens A 125: 415–421
Google ученый
Пал П., Сато К., Госальвез М.А., Тан Б., Хида Х., Шикида М. (2011) Изготовление новых микроструктур на основе ориентационно-зависимой адсорбции молекул поверхностно-активного вещества в растворе ТМАГ.Дж. Micromech Microeng 21 (1): 015008, 11pp
Google ученый
Пал П., Сато К. (2010) Методы изготовления на основе процесса влажного травления для реализации кремниевых МЭМС-структур новой формы. Microsystem Technologies 16 (7): 1165–1174
Google ученый
Gosalvez MA, Tang B, Pal P, Sato K, Kimura Y, Ishibashi K (2009) Адсорбция поверхностно-активного вещества, зависящая от ориентации и концентрации, на кремнии в водных щелочных растворах: объяснение изменений скорости травления, шероховатости и поднутрения для приложений MEMS.Дж. Micromech Microeng 19 (12): 125011, 18pp
Google ученый
Ян Г, Чан Филип Ч., Хсинг И.М., Шарма Р.К., Син JKO, Ван И (2001) Усовершенствованный раствор для травления кремнием TMAH без повреждения открытого алюминия. Актуаторы Sens A 89: 135–141
Google ученый
Chen PH, Peng HY, Hsieh CM, Chyu MK (2001) Характерное поведение водного раствора TMAH для анизотропного травления как на кремниевой подложке, так и на слое SiO 2 .Актуаторы Sens A 93 (2): 132–137
Google ученый
Tellier CR, Charbonnieras AR (2003) Характеристика анизотропного химического воздействия (hhl) кремниевых пластин в растворе TMAH 25 мас.%: Микрообработка и адекватность поверхности медленного растворения. Актуаторы Sens A 105: 62–75
Google ученый
Zhang J, Hon WC, Leung LLW, Chen KJ (2005) КМОП-совместимые методы микромашиностроения для изготовления высокопроизводительных пассивных ВЧ / СВЧ компонентов на кремниевых подложках.J Micromech Microeng 15: 328–335
Google ученый
Steinsland E, Finstad T, Hanneborg A (2000) Скорости травления (100), (111) и (110) монокристаллического кремния в TMAH, измеренные на месте с помощью лазерной интерферометрии отражения. Актуаторы Sens A 86: 73–80
Google ученый
Wu MP, Wu QH, Ko WH (1986) Исследование глубокого травления кремния с использованием этилендиамин-пирокатехин-воды.Актуаторы Sens A 9: 333–343
Google ученый
Reisman A, Berkenblit M, Chan SA, Kaufmann FB, Green DC (1979) Контролируемое травление кремния в катализированных растворах этилен-диамин-пирокатехин-вода. J Electrochem Soc: Solid-State Sci Technol 126: 1406–15
Google ученый
Керн В. (1978) Химическое травление кремния, германия, арсенида галлия и фосфида галлия.RCA Review 39: 278–307
Google ученый
Declercq MJ, Gerzberg L, Meindl JD (1975) Оптимизация раствора гидразин-вода для анизотропного травления кремния в технологии интегральных схем. J Electrochem Soc: Наука о твердом теле 122: 545–552
Google ученый
Schnakenberg U, Benecke W, Lochel B (1990) NH 4 Травитель на основе OH для микрообработки кремния.Актуаторы Sens A 23: 1031–1035
Google ученый
Clarck LD Jr, Lund JL, Edell DJ (1988) Гидроксид цезия (CsOH): полезный травитель для микрообработки кремния. В: Тех. Дайджест, IEEE Solid State Sensor and Actuator Workshop (Hilton Head Island, SC, 6-9 июня 1988 г.), стр. 5–8. http://ieeexplore.ieee.org/xpl/articleDetails.jsp?arnumber=26419
Робертсон С.В., Катехи LPB, Ребеиз Г.М. (1996) Микромашинные фильтры w-диапазона.Протоколы IEEE по теории и методам микроволнового излучения 44: 598–606
Google ученый
Blondy P, Brown AR, Cros D, Rebeiz GM (1998) Микромашинные фильтры с низкими потерями для систем связи миллиметрового диапазона. IEEE Trans Microwave Theory Tech 46: 2308–2316
Google ученый
Papapolymerou I, Drayton RF, Katehi LPB (1998) Микромашинные патч-антенны. Транзакции IEEE по антеннам и распространению информации 46: 275–83
Google ученый
Ребеиз Г. М. (2003) RF MEMS: теория, дизайн и технология. John Wiley & Sons Inc.
Burrer C, Esteve J, Lora-Tamayo E (1996) Резонансные кремниевые акселерометры в технологии объемной микрообработки – подход. J Microelectromech Syst 5: 122–130
Google ученый
Yu JC, Lan CB (2001) Системное моделирование микроакселерометра с использованием тонких пьезоэлектрических пленок. Актуаторы Sens A 88: 178–186
Google ученый
Wur DR, Davidson JL, Kang WP, Kinser DL (1995) Датчик давления из поликристаллического алмаза. J Microelectromech Syst 4: 34–41
Google ученый
Wang CC, Gogoi BP, Monk DJ, Mastrangelo CH (2000) Нечувствительные к загрязнениям дифференциальные емкостные датчики давления. J Microelectromech Syst 9: 538–543
Google ученый
Bae B, Flachsbart BR, Park K, Shannon MA (2004) Оптимизация конструкции пьезорезистивного датчика давления с учетом отношения выходного сигнала к шуму.J Micromech Microeng 14: 1597–1607
Google ученый
Yang H, Bao M, Yin H, Shen S (2002) Новый объемный микромашинный гироскоп, основанный на прямоугольной структуре пучка и массы. Актуаторы Sens A 96: 145–151
Google ученый
Van Herwaarden AW, Van Duyn DC, Van Oudheusden BW, Sarro PM (1989) Интегрированные датчики термобатареи. Актуаторы Sens A 22: 621–630
Google ученый
Sarro PM, van Hexwaarden AW, van der Vlist W (1994) Процесс изготовления кремний-кремниевой мембраны из нитрида кремния для интеллектуальных термодатчиков. Актуаторы Sens A 41–42: 666–671
Google ученый
Dillner U, Kessler E, Poser S, Baier V, Mtiller J (1997) Тепловой датчик расхода газа с низким энергопотреблением на основе термобатарей из высокоэффективных термоэлектрических материалов. Актуаторы Sens A 60: 1–4
Google ученый
Олсон Е.А., Ефремов М.Ю., Чжан М., Чжан З., Аллен Л.Х. (2003) Конструкция и работа дифференциального сканирующего нанокалориметра MEMS для высокоскоростных измерений теплоемкости ультратонких пленок. J Microelectromech Syst 12: 355–364
Google ученый
Winter W, Hohne GWH (2003) Чип-калориметр для малых образцов. Thermochimica Acta 403: 43–53
Google ученый
Zhang Y, Tadigadapa S (2004) Калориметрические биосенсоры со встроенными микрофлюидными каналами. Биосенсоры и биоэлектроника 19: 1733–1743
Google ученый
Koch M, Schabmueller CGJ, Evans AGR, Brunnschweiler A (1999) Микромашинная химическая реакционная система. Актуаторы Sens A 74: 207–210
Google ученый
Kwon JW, Kim ES (2002) Многоуровневая маршрутизация микрофлюидных каналов с защищенными выпуклыми углами.Актуаторы Sens A 97–98: 729–733
Google ученый
Pal P, Sato K (2009) Различные формы кремниевых автономных микрофлюидных каналов и микроструктур в одностадийной литографии. J Micromech Microeng 19 (5): 055003 (11pp)
Google ученый
Kwon JW, Yu H, Kim ES (2005) Методы переноса пленки и связывания для покрытия однокристальной эжекторной решетки с микроканалами и резервуарами.J Microelectromech Syst 14 (6): 1399–1408
Google ученый
Вашист С.К. (2007) Обзор микрокантилеверов для сенсорных приложений. AZoJono – Журнал. Нанотехнологии 3: 1–15
Google ученый
Wee KW, Kang GY, Park J, Kang JY, Yoon DS, Parkb JH, Kim TS (2005) Новое электрическое обнаружение белков-маркеров болезней без меток с использованием пьезорезистивных самочувствительных микрокантилеверов.Биосенсоры и биоэлектроника 20: 1932–1938
Google ученый
Lee JH, Hwang KS, Park J, Yoon KH, Yoon DS, Kim TS (2005) Иммуноанализ простатоспецифического антигена (PSA) с использованием резонансного сдвига частоты пьезоэлектрического наномеханического микрокантилевера. Биосенсоры и биоэлектроника 20: 2157–62
Google ученый
Battiston FM, Ramseyer JP, Lang HP, Baller MK, Gerber C, Gimzewski JK, Meyer E, Guntherodt HJ (2001) Химический датчик на основе микроволоконного массива консолей с одновременным считыванием резонансной частоты и изгиба.Актуаторы Sens B 77: 122–131
Google ученый
Neuzil P, Nagarajan R (2006) Формирование острых острий AFM путем одноступенчатого травления. J Micromech Microeng 16: 1298–1300
Google ученый
Берт Д.П., Добсон П.С., Дональдсон Л., Уивер Дж.М.Р. (2008) Простой метод высокопрочного изготовления острых силиконовых наконечников. Микроэлектронная инженерия 85: 625–630
Google ученый
Han J, Lu S, Li Q, Li X, Wang J (2009) Анизотропное влажное травление кремниевых наконечников с малым углом раскрытия в растворе KOH с добавками I2 / KI. Актуаторы Sens A 152: 75–79
Google ученый
Пал П., Сингх С.С. (2013) Новая модель характеристик травления углов, образованных плоскостями Si {111} на поверхности пластины Si {110}. Инженерное дело 5 (11): 1–8
Google ученый
Trieu HK, Mokwa W (1998) Обобщенная модель, описывающая подрезание угла путем экспериментального анализа TMAH / IPA. J Micromech Microeng 8: 80–83
Google ученый
Chahoud M, Wehmann HH, Schlachetzki A (1998) Моделирование травления выпуклых и смешанных структур InP и Si. Актуаторы Sens A 69: 251–258
Google ученый
Schroder H, Obermeier E (2000) Новая модель для вырезания выпуклых углов Si {100} при анизотропном травлении KOH.J Micromech Microeng 10: 163–170
Google ученый
Шикида М., Нанбара К., Коидзуми Т., Сасаки Х., Сато К., Одагаки М., Андо М., Фурута С., Асауми К. (2000) Модель, объясняющая явление поднутрения угла маски при анизотропном травлении кремния: седло точка на диаграмме скорости травления. Актуаторы Sens A 97–98: 758–63
Google ученый
Chang Chien WT, Chang CO, Lo YC, Li ZW, Chou CS (2005) Об определении индексов Миллера плоскостей поднутрения выпуклых углов Si {100}.J Micromech Microeng 15: 833–842
Google ученый
Merlos A, Acero MC, Bao MH, Bausells J, Esteve J (1992) Исследование характеристик поднутрения в системе TMAH: IPA. J Micromech Microeng 2: 181–183
Google ученый
Dong W, Zhang X, Liu C, Li M, Xu B, Chen W. (2004) Механизм для выпуклой подрезки углов кремния (110) в КОН. Microelectronics J 35: 417–419
Google ученый
Пал П., Сингх С.С. (2013) Простая и надежная модель, объясняющая подрезание выпуклых углов при микрообработке мокрых массивов. Письма о микро- и наносистемах 1 (1): 1–6
MathSciNet Google ученый
Бин К.Э., Рунян В.Р. (1977) Диэлектрическая изоляция: комплексные, текущие и будущие. J Electrochem Soc 124: 5C – 12C
Google ученый
Abu-Zeid M (1984) Подрезка угла в анизотропно протравленных изоляционных контурах.J Electrochem Soc 131: 2138–2142
Google ученый
Wu XP, Ko WH (1989) Компенсирующая подрезка углов при анизотропном травлении кремния (100). Актуаторы Sens A 18: 207–215
Google ученый
Пуэрс Б., Сансен В. (1990) Компенсационные структуры для микрообработки выпуклых углов в кремнии. Актуаторы Sens A 23: 1036–1041
Google ученый
Майер Г.К., Офферинс Х.Л., Сандмайер Х., Кул К. (1990) Изготовление невытянутых выпуклых углов при анизотропном травлении кремния (100) в водном растворе КОН с учетом новых микромеханических элементов. J Electrochem Soc 137: 3947–3951
Google ученый
Offereins HL, Kühl K, Sandmaier H (1991) Методы изготовления выпуклых углов при анизотропном травлении кремния (100) в водном растворе КОН. Актуаторы Sens A 25: 9–13
Google ученый
Sandmaier H, Offereins HL, Kuhl K, Lang W. (1991) Методы компенсации углов при анизотропном травлении (100) -кремния с использованием водного КОН. In: 6 th International Conference on Solid State Sensors and Actuators (Transducers 91, San Francisco, CA, 24-27 июня 1991), стр. 456–459. http://ieeexplore.ieee.org/xpl/login.jsp?tp=&arnumber=148910&url=http%3A%2F%2Fieeexplore.ieee.org%2Fiel2%2F505%2F3940%2F00148910.pdf%3Farnumber%3D148910
Hui WC (1991) Методика защиты углов микросхем при влажном химическом травлении кремниевых пластин.Технический отчет Национальная лаборатория Лоуренса Ливермора, Калифорния (США)
Google ученый
Offereins HL, Sandmaier H, Marusczyk K, Kuhl K, Plettner A (1992) Компенсирующая угловая подрезка кремния (100) в КОН. Датчики и материалы 3: 127–144
Google ученый
Bao M, Chr B, Esteve J, Bausells J, Marco S (1993) Травление переднего контроля полос <110> для компенсации углов.Актуаторы Sens A 37–38: 727–732
Google ученый
Scheibe C, Obermeier E (1995) Компенсирующая подрезка углов при анизотропном травлении кремния (100) для разделения чипа. J Micromech Microeng 5: 109–111
Google ученый
Кампен Р.П., Вольфенбюттель Р.Ф. (1995) Влияние <110> -ориентированных угловых компенсационных структур на качество мембраны и целостность выпуклых углов в кремнии (100) с использованием водного КОН.J Micromech Microeng 5: 91–94
Google ученый
Zhang Q, Liu L, Li Z (1996) Новый подход к компенсации выпуклых углов при анизотропном травлении Si (100) в КОН. Актуаторы Sens A 56: 251–254
Google ученый
Enoksson P (1997) Новая структура для компенсации углов при анизотропном травлении КОН. J Micromech Microeng 7: 141–144
Google ученый
Chung CK, Lee CC, Wu CY (1998) Новый подход к компенсации углов многоступенчатой террасной структуры Si (100) для микролинз. Летняя тематическая встреча IEEE / LEOS, Монтерей, Калифорния
Google ученый
Long M K, Burdick J W, Antonsson E K (1999) Расчет компенсационных структур для анизотропного травления. В: Технические материалы Международной конференции 1999 г. по моделированию и моделированию микросистем (MSM 99).
Ma L (2001) Надежная компоновка маски и синтез процесса в микро-электромеханических системах (MEMS) с использованием генетических алгоритмов. Кандидат наук. Диссертация Калифорнийский технологический институт Пасадена, Калифорния
Google ученый
Tellier C (2003) САПР-дизайн шаблонов компенсации маски. Proc IEEE Sensors 1: 517–522
Google ученый
Wacogne B, Sadani Z, Gharbi T (2004) Компенсационные структуры для V-образных канавок, соединенных с квадратными отверстиями в кремнии с KOH-травлением (100): теория, моделирование и эксперименты.Актуаторы Sens A 112: 328–339
Google ученый
Fan W, Zhang D (2006) Простой подход к компенсации выпуклых углов при анизотропном травлении KOH на кремниевой пластине (100). J Micromech Microeng 16: 1951–1957
Google ученый
Mukhiya R, Bagolini A, Margesin B, Zen M, Kal S (2006) <100> компенсация угла стержня для CMOS-совместимого анизотропного травления TMAH.J Micromech Microeng 16: 2458–2462
Google ученый
Пал П., Сато К., Чандра С. (2007) Методы изготовления выпуклых углов в (100) -кремниевой пластине с использованием объемной микрообработки: обзор. J Micromech Microeng 17: R1 – R23
Google ученый
Бисвас К., Дас С., Кал С. (2006) Анализ и предотвращение подрезания выпуклых углов в объемных микроструктурах кремния, подвергнутых механической обработке.Microelectronics J 37: 765–769
Google ученый
Pal P, Chandra S (2004) Утопленные микроструктуры с идеально выпуклыми углами для акселерометров. Sensor Letters 2: 226–231
Google ученый
Pal P, Chandra S (2004) Новый процесс для реализации идеальных выпуклых углов при объемной микрообработке. J Micromech Microeng 14: 1416–1420
Google ученый
Кавакацу Х., Сая Д., Като А., Фукусима К., Тошиёси Х., Фудзита Х. (2002) Миллионы кантилеверов для атомно-силовой микроскопии. Обзор научных инструментов 73 (3): 1188–1192
Google ученый
Chu HY, Fang W (2004) Компенсация вертикальных выпуклых углов и защита не {111} кристаллических плоскостей для процесса влажной анизотропной объемной микрообработки. J Micromech Microeng 14: 806–813
Google ученый
Jia C, Dong W, Liu C, Zhang X, Zhou J, Zhong Z, Xue H, Zang H, Xu B, Chen W (2006) Подрезание выпуклых углов и компенсация ромбов в KOH с раствором IPA и без него (110) кремний. Microelectronics J 37: 1297–1301
Google ученый
Ким Б., Чо Д.Д. (1998) Водное травление кремния КОН (110) Характеристики травления и методы компенсации выпуклых углов. J Electrochem Soc 145: 2499–508
Google ученый
Ciarlo DR (1987) Угловые компенсационные структуры для кремния с ориентацией (110). Proc. IEEE Micro Robots and Teleoperators Workshop, Хианнис, Массачусетс, США
Google ученый
Wacogne B, Zeggari R, Sadani Z, Gharbi T (2006) Очень простой метод компенсации изогнутых V-образных канавок в кремнии, протравленном KOH (100), когда требуются тонкие структуры или глубокое травление. Актуаторы Sens A 126: 264–269
Google ученый
Куммамуру Р.К., Ху Л., Кук Л., Ефремов М.Ю., Олсон Э.А., Аллен Л.Х. (2008) Самовыравнивающаяся теневая маска для непосредственного нанесения на мембрану или полость. J Micromech Microeng 18: 0
(9pp)
Google ученый
Пал П., Сато К., Шикида М., Госальвез М.А. (2009) Изучение структур с компенсацией углов и изготовление различных форм структур MEMS в чистом TMAH с добавлением поверхностно-активного вещества. Актуаторы Sens A 154: 192–203
Google ученый
Yu JC (2011) Компенсация выпуклого угла для компактной сейсмической массы с высоким аспектным отношением с использованием анизотропного влажного травления кремния (100). В: Симпозиум по проектированию, тестированию, интеграции и упаковке MEMS / MOEMS (DTIP-11, Экс-ан-Прованс, Франция, 11-13 мая 2011 г.), стр. 197–199. http://ieeexplore.ieee.org/xpl/login.jsp?tp=&arnumber=6107993&url=http%3A%2F%2Fieeexplore.ieee.org%2Fiel5%2F60
%2F6107967%2F06107993.pdf%3Farn99%3DF%6107967%3DMukhiya R, Bagolini A, Bhattacharya TK, Lorenzelli L, Zen M (2011) Экспериментальное исследование и анализ структур компенсации углов для CMOS-совместимой объемной микрообработки с использованием 25 мас.% TMAH.Microelectronics J 42: 127–134
Google ученый
Bagolini A, Faes A, Decarli M (2010) Влияние потенциала травления на анизотропное травление выпуклых углов в растворе TMAH. J Microelectromech Syst 19 (5): 1254–1259
Google ученый
Smiljanic MM, Jovic V, Lazic Z (2012) Метод компенсации выпуклых углов без маски на кремниевой подложке (100) в 25% -ном водном растворе TMAH.J Micromech Microeng 22: 115011 (11pp)
Google ученый
Шаян М., Арезоо Б., Ванини А.С., Котамджани А.А. (2011) Моделирование поднутрения кремния (100) и расчет структур с компенсацией углов в процессе влажного анизотропного травления. Proc. IMechE Часть B: J. Машиностроение 225: 1041–1049. http://pib.sagepub.com/content/225/7/1041.abstract
Апаниус М., Каул Б., Абрамсона А.Р. (2007) Изготовление кремниевой теневой маски для узорчатого осаждения металла с микромасштабными размерами с использованием новой схемы компенсации углов.Актуаторы Sens A 140: 168–175
Google ученый
Giousouf M, Assmus F, Kuck H (1999) Структурирование выпуклых углов с использованием процесса повторного окисления в резонаторе камертона, сделанном из (110) -кремния. Датчики и исполнительные механизмы A 76: 416–424
Google ученый
Pal P, Gosalvez MA, Sato K, Hida H, Xing Y (2014) Анизотропное травление на Si {110}: эксперимент и моделирование формирования микроструктур с выпуклыми углами.J Micromech Microeng 24: 125001 (25pp)
Google ученый
Cho WJ, Chin WK, Kuo CT (2004) Влияние спиртовых замедлителей на анизотропное травление кремния в водных растворах гидроксида калия. Актуаторы Sens A 116: 357–368
Google ученый
Zubel I, Kramkowska M (2004) Скорость травления и морфология поверхностей кремния (hkl), протравленных в KOH и KOH, насыщенных растворами изопропанола.Актуаторы Sens A 115: 549–556
Google ученый
Philipsen HGG, Kelly JJ (2009) Влияние химических добавок на поверхностную реакционную способность Si в растворе КОН. Electrochimica Acta 54: 3526–3531
Google ученый
Zubel I, Kramkowska M (2001) Влияние изопропилового спирта на скорость травления и шероховатость поверхности (100) Si, протравленной в растворах KOH и TMAH.Датчики и исполнительные механизмы A 93: 138-147. http://www.sciencedirect.com/science/article/pii/S04701006483
Зубель И., Крамковская М. (2002) Влияние спиртовых добавок на характеристики травления в растворах КОН. Актуаторы Sens A 101: 255–261
Google ученый
Pal P, Gosalvez MA, Sato K (2010) Микрообработка кремния на основе гидроксида тетраметиламмония с добавлением поверхностно-активного вещества: механизм травления и расширенное применение.Japan J Appl Phys 49: 056702 (9pp)
Google ученый
Госальвез М.А., Пал П., Танг Б., Сато К. (2010) Атомистический механизм макроскопических эффектов, вызванных небольшими добавками поверхностно-активных веществ в щелочные травильные растворы. Актуаторы Sens A 157: 91–95
Google ученый
Tang B, Pal P, Gosalvez MA, Shikida M, Sato K, Amakawa H, Itoh S (2009) Эллипсометрическое исследование толщины адсорбированного поверхностно-активного вещества на Si {110} и Si {100} и влияние предварительно адсорбированный слой поверхностно-активного вещества на характеристики травления в TMAH.Актуаторы Sens A 156: 334–341
Google ученый
Pal P, Sato K, Gosalvez MA, Kimura Y, Ishibashi K, Niwano M, Hida H, Tang B, Itoh S (2009) Адсорбция поверхностно-активного вещества на поверхности монокристаллического кремния в растворе TMAH: адсорбция в зависимости от ориентации обнаруживается с помощью инфракрасной спектроскопии на месте. J Microelectromech Syst 18: 1345–1356
Google ученый
Ян Ч.Р., Чен П.Й., Ян Ч.Х., Чиу Ю.К., Ли Р.Т. (2005) Влияние различных ионных поверхностно-активных веществ на свойства анизотропного травления кремния в растворах КОН и ТМАГ.Актуаторы Sens A 119: 271–281
Google ученый
Yang CR, Yang CH, Chen PY (2005) Исследование характеристик анизотропного травления кремния в водных растворах гидроксида тетраметиламмония с добавлением различных поверхностно-активных веществ. J Micromech Microeng 15: 2028–2037
Google ученый
Сато К., Учикава Д., Шикида М. (2001) Изменение свойств травления монокристаллического кремния, зависящих от ориентации, вызванное поверхностно-активным веществом, добавленным к раствору ТМАГ.Sens Mater 13: 285–291
Google ученый
Сарро П.М., Брида Д., ван дер Влист В., Брида С. (2000) Влияние поверхностно-активного вещества на качество поверхности кремниевых микроструктур, протравленных в насыщенных растворах TMAHW. Датчики Актуаторы A 85: 340–345
Google ученый
Sekimura M (1999) Анизотропное травление раствора ТМАГ с добавлением поверхностно-активного вещества. В: Proc. 12-я Международная конференция IEEE по микроэлектромеханическим системам.(MEMS-99 Орландо, Флорида, 17–21 января 1999 г.), стр. 650–655. http://ieeexplore.ieee.org/xpl/login.jsp?tp=&arnumber=746904&url=http%3A%2F%2Fieeexplore.ieee.org%2Fxpls%2Fabs_all.jsp%3Farnumber%3D746904
Ян С.Р., Чен П.Й., Чиу Ю.К., Ли Р.Т. (2005) Влияние механического перемешивания и добавки поверхностно-активного вещества на анизотропное травление кремния в щелочном растворе КОН. Актуаторы Sens A 119: 263–270
Google ученый
Resnik D, Vrtacnik D, Aljancic U, Mozek M, Amon S (2005) Роль поверхностно-активного вещества Тритон в анизотропном травлении отражающих плоскостей {110} на кремнии (100). J Micromech Microeng 15: 1174–1183
Google ученый
Xu YW, Michael A, Kwok CY (2011) Формирование сверхгладкого микрозеркала под углом 45 ° на кремнии (100) с низкой концентрацией TMAH и поверхностно-активным веществом: методы увеличения истинно 45 ° части. Актуаторы Sens A 166: 164–71
Google ученый
Pal P, Sato K, Gosalvez MA, Shikida M (2007) Изучение подрезания закругленных вогнутых и острых краев выпуклых углов в КМОП-совместимых анизотропных травителях. J Micromech Microeng 17: 2299–2307
Google ученый
Zubel I, Kramkowska M, Rola K (2012) Анизотропное травление кремния в растворах TMAH, содержащих спиртовые и поверхностно-активные добавки. Актуаторы Sens A 178: 126–135
Google ученый
Rola KP, Zubel I (2013) Поверхностно-активное вещество тритон как добавка к травителю кремния KOH. J Microelectromech Syst 22: 1373–1382
Google ученый
Танака Х., Умэки Н., Сато К. (2013) Идеальная адсорбция поверхностно-активного вещества на уровне частей на миллиард в 5% водном растворе КОН на поверхности кремния, изменяющая свойства анизотропного травления. In: 17 th International Conference on Solid State Sensors, Actuators and Microsystems (Transducers & Eurosensors XXVII, Barcelona, Spain, 16-20 июня 2013) pp 1978–1981.http://ieeexplore.ieee.org/xpl/articleDetails.jsp?arnumber=6627183
Винд Р.А., Хайнс М.А. (2000) Макроскопическая анизотропия травления и микроскопические механизмы реакции: микромашинная структура для быстрого анализа анизотропии травителя. Наука о поверхности 460: 21–38
Google ученый
Винд Р.А., Джонс Х., Литтл М.Дж., Хайнс М.А. (2002) Химическая кинетика с ориентационным разрешением: использование микротехнологий для разгадки сложной химии травления KOH / Si.J Phys Chem B 106: 1557–1569
Google ученый
Госальвез М.А., Пал П., Феррандо Н., Хида Х., Сато К. (2011) Экспериментальное получение полного трехмерного распределения скорости травления Si в анизотропных травителях на основе вертикально подвергнутых микромеханической обработке образцов колес вагонов. J Micromech Microeng 21: 125007 (14 страниц)
Google ученый
Госальвез М.А., Пал П., Феррандо Н., Сато К. (2011) Оценка надежности полного трехмерного распределения скорости травления Si в анизотропных травителях на основе вертикально обработанных микромеханических образцов колес вагонов.J Micromech Microeng 21: 125008 (12 стр.)
Google ученый
Госальвез М.А., Пал П., Сато К. (2011) Восстановление трехмерного распределения скорости травления кремния в анизотропных травителях с использованием данных с вицинальных поверхностей {100}, {110} и {111}. J Micromech Microeng 21: 105018 (17pp)
Google ученый
Джеймс Т.Д., Пэриш Г., Винчестер К.Дж., Маска Калифорния (2006) Метод кристаллографического выравнивания в кремнии для изготовления глубоких и длинных микроканалов.J Micromech Microeng 16: 2177–2182
Google ученый
Chang WH, Huang YC (2005) Новый шаблон предварительного травления для определения кристаллографической ориентации <110> на кремниевых пластинах (100) и (110). Микросистемные технологии 11: 117–128
Google ученый
Ensell G (1996) Выравнивание шаблона маски по ориентации кристалла. Актуаторы Sens A 53: 345–8
Google ученый
Vangbo M, Bäcklund Y (1996) Точное выравнивание маски по кристаллографической ориентации кремниевых пластин с использованием влажного анизотропного травления. J Micromech Microeng 6: 279–284
Google ученый
Lai JM, Chieng WH, Huang YC (1998) Точное выравнивание травления маски относительно ориентации кристалла. J Micromech Microeng 8: 327–329
Google ученый
Ценг Ф.Г., Чанг К.С. (2003) Точные определения [100] ориентации кристаллов на кремниевых пластинах с ориентацией <110>. J Micromech Microeng 13: 47–52
Google ученый
Pal P, Chandra S (2004) Объемные микромашинные структуры внутри анизотропно протравленных полостей. Smart Mater Struct 13: 1424–1429
Google ученый
Чандра С., Сингх Дж., Чанд А. (1997) Дизайн и разработка микроструктур для приложений МЭМС.Proc SPIE 3226: 22–30
Google ученый
Абединов Н., Грабец П., Готсальк Т., Иванов Т.З., Фойгт Дж., Ренджлоу И.В. (2001) Микромашинная пьезорезистивная консольная решетка со встроенным резистивным микронагревателем для калориметрии и масс-детектирования. J Vac Sci Technol A 19: 2884–2888
Google ученый
Госальвез М.А. (2003) Атомистическое моделирование анизотропного травления кристаллического кремния.Кандидатская диссертация, диссертация 123, диссертации лаборатории физики Хельсинкского технологического университета.
Jaccodine RJ (1962) Использование модифицированных теорем о свободной энергии для предсказания равновесных форм роста и травления. J Appl Phys 33: 2643–2647
Google ученый
Pal P, Haldar S, Singh SS, Ashok A, Xing Y, Sato K (2014) Подробное исследование и объяснение появления различных профилей поднутрения в KOH и TMAH.J Micromech Microeng 24: 0
(9pp)Google ученый
Кучоуков В.Г., Моллингер Дж. Р., Босше А. (1999) Новый метод формования фоторезиста на пластинах со сквозным отверстием. В: Proc. 13-я Европейская конф. на твердотельных преобразователях (Евросенсоры), стр. 256–72
Крейвен Д. (1996) Проблемы фотолитографии для микромеханической промышленности, Proc. SPIE 2884, 16 th Ежегодный симпозиум BACUS по технологии и управлению фотошаблонами, 498 (декабрь.27, 1996), с. 1–10. http://spie.org/Publications/Proceedings/Paper/10.1117/12.262838
Venstra WJ, Spronck JW, Sarro PM, Eijk JV (2009) Фотолитография на объемных микромеханически обработанных подложках. J Micromech Microeng 19: 055005 (6pp)
Google ученый
Luxbacher T, Mirza A (1999) Распылительное покрытие для МЭМС, межкомпонентных соединений и сложных упаковочных приложений. Датчики 16: 61–64
Google ученый
Сингх В.К., Сасаки М., Хейн К., Эсаши М. (2004) Условия потока при нанесении напыляемого резиста и характеристики формирования рисунка для трехмерной фотолитографии над глубокими структурами. Japan J Appl Phys 43: 2387–2391
Google ученый
Linder S, Baltes H, Gnaedinger F, Doering E (1996) Фотолитография в канавках с анизотропным травлением. В: Proc. 9 th Ежегодный международный семинар по микроэлектромеханическим системам (MEMS-96, 11-15 февраля 1996 г.), стр. 38-43.http://ieeexplore.ieee.org/xpl/login.jsp?tp=&arnumber=4&url=http%3A%2F%2Fieeexplore.ieee.org%2Fxpls%2Fabs_all.jsp%3Farnumber%3D4
Kersten P, Bouwstra S, Petersen JW (1995) Фотолитография на микромашинных 3-D поверхностях с использованием электроосажденного фоторезиста. Актуаторы Sens A 51: 51–54
Google ученый
Heschel M, Bouwstra S (1998) Конформное покрытие фоторезистом острых углов анизотропно протравленных сквозных отверстий в кремнии.Актуаторы Sens A 70: 75–80
Google ученый
Pal P, Sato K (2009) Сложные трехмерные структуры в Si {100} с использованием влажной объемной микрообработки. Дж. Micromech Microeng 19: 105008 (9pp)
Google ученый
Appels JA, Kooi E, Paffen MM, Schatorje JJH, Verkuylen WHCG (1970) Локальное окисление кремния и его применение в технологии полупроводниковых устройств.Philips Res Repts 25: 118–132
Google ученый
Loewenstein LM, Tipton CM (1991) Химическое травление термически окисленного нитрида кремния: сравнение методов влажного и сухого травления. J Electrochem Soc 138: 1389–1394
Google ученый
% PDF-1.7 % 2 0 obj > эндобдж 4 0 obj > транслировать application / pdfuuid: 4a1a5922-1162-4400-b7c4-49aad5c87b45uuid: b04e702a-eead-4dff-bdbf-1c922dc7ace52016-06-27T12: 12: 44 + 02: 00PDF-XChange Editor 5.5.316.12016-06-27T12: 12: 53 + 02: 00PDF-XChange PDF Core API (5.5.316.1) конечный поток эндобдж 3 0 obj > транслировать xQo6) ˒3Crx ( NFiq ً7 XowIѸČ | 3! E |, A> LU] e1Cq = 2y “.ocF ſ / ί ǿu +? ~ D
Ixodes scapularis Белки слюны клеща, последовательно секретируемые каждые 24 часа во время кормления кровью
Цитата: Kim TK, Tirloni L, Pinto AFM, Moresco J, Yates JR III, da Silva Vaz I Jr, et al. (2016) Ixodes scapularis Белки слюны клещей последовательно секретируются каждые 24 часа во время кормления кровью. PLoS Negl Trop Dis 10 (1): e0004323. https://doi.org/10.1371/journal.pntd.0004323
Редактор: Роэль Рамос Дингласан, Школа общественного здравоохранения Bloomberg Джонса Хопкинса, США
Поступила: 30 сентября 2015 г .; Принята к печати: 2 декабря 2015 г .; Опубликовано: 11 января 2016 г.
Авторские права: © 2016 Kim et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника
Доступность данных: Данные масс-спектрометрии, протеомики были депонированы в Консорциум ProteomeXchange через партнерский репозиторий PRIDE с идентификатором набора данных PXD003214.
Финансирование: Это исследование было поддержано грантами Национального института здравоохранения (AI081093, AI0
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Клещи превосходят всех членистоногих в передаче большего числа патогенов, включая грибы, вирусы, бактерии и простейшие [1, 2].В животноводстве клещи и клещевые болезни (TBD) вызывают ежегодные убытки в миллиарды долларов США во всем мире [3, 4]. В последнее время клещи привлекли внимание в политике общественного здравоохранения в связи с недавней публикацией, в которой пропагандировалось решение One Health, в котором 17 случаев TBD у человека были включены в число источников проблем со здоровьем [5]. Ixodes scapularis , широко известный как олень, или черноногий клещ, является одним из наиболее важных с медицинской точки зрения видов клещей и передает 5 из 14 возбудителей TBD человека в США: Borrelia burgdorferi [6], Anaplasma phagocytophilum [7] ], Borrelia miyamotoi [8], Babesia microti [9] и болезнь, вызванная вирусом Повассан [10].Точно так же близкие родственники этого клеща, включая I . pacificus на западном побережье США и I . ricinus в Европе являются переносчиками важных возбудителей TBD человека, включая B . burgdorgferi , B . miyamotoi и A . phagocytophilum [11]. Исходя из этого, I . scapularis Геном был секвенирован [12, 13], и эти данные предоставили возможность для углубленных исследований биологических адаптаций, которые делают клещей успешными переносчиками патогенов.Эти данные были постулированы для облегчения исследований, которые выявят слабые места, которые могут быть использованы для разработки новых методов борьбы с клещами [13].
В отсутствие эффективных вакцин против TBD, борьба с клещами с помощью акарицидов остается наиболее надежным методом [14, 15]. Несмотря на свою эффективность в краткосрочной перспективе, ограничения химических акарицидов, такие как отбор устойчивых популяций клещей, затраты на разработку новых акарицидов, загрязнение окружающей среды и пищевых продуктов, вызвали необходимость поиска альтернативных методов борьбы с клещами [4, 16, 17].Иммунизация животных считается устойчивым альтернативным методом борьбы с клещами [18]. Основным ограничением глобального принятия противоклещевых вакцин в качестве альтернативного метода борьбы с клещами является наличие эффективных целевых антигенов. Мы заинтересованы в понимании физиологии кормления клещей как средства идентификации физиологически важных белков, которые могут быть нацелены на разработку вакцины против клещей.
Клещи совершают кормление, разрывая сосудистую сеть окружающей ткани хозяина и всасывая кровь, которая кровоточит в пораженное место кормления [19–21].Этот деструктивный стиль питания запускает защитные механизмы хозяина, такие как гемостаз, воспаление и иммунные реакции. Однако клещи противодействуют защитным механизмам хозяина, секретируя фармакологически активные молекулы в слюну, чтобы модулировать защиту хозяина [19, 20, 22–24]. Помимо получения кровяной муки, белки слюны клещей также участвуют в передаче и приобретении возбудителей TBD [25]. Сообщения о снижении передачи патогенов животным, неоднократно заражавшимся клещами, у которых развилась устойчивость к питанию клещами [26–29], подтверждают важность белков слюны клещей в компетентности переносчиков клещей.Таким образом, идентификация белков слюны клещей станет основой для разработки новых методов, препятствующих питанию клещей и предотвращения передачи патогенов.
С появлением технологий секвенирования следующего поколения (NGS) были описаны транскриптомы слюнных желез клещей [23, 30–39]. Однако основным ограничением этих данных является то, что они не информируют о том, какие транскрипты, кодирующие белки, секретируются в слюне клещей. В интересном подходе к идентификации секретируемых белков слюны клещей (TSP) Radulovic et.др., [40] и Lewis et. др. [41] использовали антитела к 24–48-часовым белкам слюны клещей [42, 43] для иммуноскрининга библиотек экспрессии кДНК фагового дисплея для идентификации 24–48 часов Amblyomma americanum и 24 часов I . scapularis иммуногенные белки слюны клещей. Аналогичные подходы иммуноскрининга были использованы для идентификации иммунодоминанта I . scapularis белки слюны клеща [44–47]. В соответствующем исследовании слюна I . scapularis анализировали деградацией по Эдману, идентифицировав 15 белков [48].Недавно белки в слюне иксодовых клещей от обильного кормления Rhipicephalus sanguineus [49], частичного и обильного кормления Rhipicephalus microplus [50], трех- и пятидневного кормления Dermacentor andersoni [51] и взрослых особей и нимфы , вскармливаемых повторно. Haemaphysalis longicornis [52]. У аргасидных клещей в ходе единственного исследования были идентифицированы белки слюны у клещей Ornithodoros moubata , которых дважды кормили, с слюной, собранной через 4 месяца после кормления [53].В то время как исследования, рассмотренные здесь, идентифицировали белки в слюне клещей в один или два момента времени кормления, в этом исследовании были описаны белки I . scapularis клещи, вероятно, вводят животным инъекции каждые 24 часа в течение первых пяти дней кормления и ближе к концу процесса кормления клещей. Каталог I . scapularis белки слюны клеща в этом исследовании дают подробный обзор семейств белков и / или молекулярных систем, которые задействованы в I . scapularis тик и хост-интерфейс.
Материалы и методы
Заявление о соблюдении этических норм
Все эксперименты проводились в соответствии с протоколом использования животных, утвержденным Комитетом по уходу и использованию животных (IACUC) Техасского университета A&M (AUP 2011–207 и 2011–189), который отвечает всем федеральным требованиям, как определено в Законе о благополучии животных (AWA). ), Политику службы общественного здравоохранения (PHS) и гуманный уход и использование лабораторных животных.
Сбор клещей и слюны
Я . scapularis клещей были приобретены в центре по выращиванию клещей в Университете штата Оклахома (Стиллуотер, штат Оклахома, США). Перед кормлением кроликов самок клещей спаривали с самцами для спаривания. Клещи считались спарившимися после отделения самцов от самок. Обычно клещей кормили кроликами, как описано ранее [43]. Вязка I . scapularis клещей были ограничены питанием на внешней части уха новозеландских кроликов с помощью ортопедического трикотажа, приклеенного клеем Kamar (Kamar Products Inc., Зайонсвилл, Индиана, США). Всего 84 взрослых I . scapularis клещей (42 на ухо) помещали в устройство для удержания клещей на трех кроликах и позволяли им прикрепиться.
Для сбора слюны клещей самки клещей частично кормили в течение 24 часов (n = 43 клещей), 48 часов (n = 40 клещей), 72 часов (n = 40 клещей), 96 часов (n = 40 клещей), 120 часов ( n = 40 клещей), а также явно полностью накормленных, но не отделившихся от хозяина (BD, n = 8 клещей) и спонтанно отделившихся клещей (SD, n = 6 клещей) промывали водой Milli-Q и сушили на бумажном полотенце. .Промытых клещей помещали дорсальной стороной вниз на двусторонний скотч на предметное стекло. Слюноотделение вызывали инъекцией 1–3 мкл 2% гидрохлорида пилокарпина в фосфатно-солевом буфере (PBS, pH 7,4) на вентральную сторону, прилегающую к тазику четвертой ноги, с помощью иглы со скошенным углом 34 размера / 0,5 дюйма / 45 ° на модели. Шприц 701 Hamilton (Hamilton Company, Рино, Невада, США). Впоследствии слюну собирали каждые 15–30 минут с помощью шприца Гамильтона в течение примерно 4 часов при комнатной температуре.
Расщепление белков и подготовка проб
Слюна I . scapularis клещей (по меньшей мере 2 мкг общего белка на цикл X3) для каждой конкретной точки времени кормления переваривали в растворе с трипсином. Слюну разбавляли 8 М мочевиной / 0,1 М трис, pH 8,5, восстанавливали 5 мМ трис (2-карбоксиэтил) фосфин гидрохлорид (TCEP, Sigma-Aldrich, Сент-Луис, Миссури, США) и алкилировали 25 мМ йодацеамидом (Sigma- Олдрич). Белки переваривали в течение ночи при 37 ° C в 2 М мочевине / 0,1 М Tris pH 8,5, 1 мМ CaCl 2 с трипсином (Promega, Мэдисон, Висконсин, США) с конечным соотношением 1:20 (фермент: субстрат).Реакции переваривания в конечной концентрации 0,15 мкг / мл гасили муравьиной кислотой (конечная концентрация 5%) и центрифугировали для удаления остатков.
Предколонки и аналитические колонки
Предварительные колонки с обращенной фазой готовили, сначала создавая фритту Kasil на одном конце деактивированного капилляра с внутренним диаметром 250 мкм и внешним диаметром 360 мкм (Agilent Technologies, Санта-Клара, Калифорния, США). Фритты Kasil получали путем погружения капилляра длиной 20 см в 300 мкл Kasil 1624 (PQ Corporation, Малверн, Пенсильвания, США) и 100 мкл раствора формамида, отверждения при 100 ° C в течение 3 часов и регулирования длины.Предварительные колонки были набиты на месте (лаборатория Джона Ятса III, Исследовательский институт Скриппса, Ла-Хойя, Калифорния, США) 2 см 5 мкм частицами ODS-AQ C18 (YMC America, Inc., Аллентаун, Пенсильвания, США). из суспензий частиц в метаноле. Аналитические колонки с обращенной фазой были изготовлены путем вытягивания капилляра из диоксида кремния с внутренним диаметром 100 мкм и внешним диаметром 360 мкм (Molex Polymicro Technologies, Остин, Техас, США) к наконечнику с внутренним диаметром 5 мкм. Тот же упаковочный материал был упакован до 20 см непосредственно за вытянутым наконечником. Предварительные колонки с обращенной фазой и аналитические колонки были соединены с использованием объединения нулевого мертвого объема (IDEX Corp., Upchurch Scientific, Ок-Харбор, Вашингтон, США).
ЖХ-МС / МС
Пептидные смеси анализировали с помощью масс-спектрометрии жидкостной хроматографии с нанопотоком с использованием масс-спектрометра Easy NanoLC II и Q Exactive (Thermo Scientific, Waltham, MA, USA). Пептиды, элюированные из аналитической колонки, распыляли электрораспылением непосредственно в масс-спектрометр. Буфер A и B состоял из 5% ацетонитрила / 0,1% муравьиной кислоты и 80% ацетонитрила / 0,1% муравьиной кислоты соответственно. Скорость потока была установлена на 400 нл / мин.Время кормления Образцы слюны (1,5 мкг на инъекцию) разделяли за 155-минутные хроматографические прогоны следующим образом: 1–10% градиент буфера B за 10 минут, 10–40% буфера B за 100 минут, 40–50% буфера. B через 10 мин и 50–90% буфера B через 10 мин. Колонку выдерживали при 90% буфера B в течение 10 минут, уменьшали до 1% буфера B и повторно уравновешивали перед следующей инъекцией.
Масс-спектрометр работал в режиме зависимости от данных, собирая полное сканирование МС от 400 до 1200 m / z при разрешении 70000 и цели AGC 1 x 10 6 .10 наиболее распространенных ионов на сканирование были выбраны для МС / МС с разрешением 17 500 и мишенью АРУ 2 x 10 5 и коэффициентом недостаточного заполнения 0,1%. Максимальное время заполнения составляло 20 и 120 мс для сканирования MS и MS / MS, соответственно, с динамическим исключением 15 с. Нормализованная энергия столкновения была установлена на 25. Данные масс-спектрометрической протеомики были депонированы в ProteomeXchange Consortium [54] через репозиторий партнеров PRIDE с идентификатором набора данных PXD003214.
Анализ данных
Тандемные масс-спектры были извлечены из файлов Thermo RAW с использованием RawExtract 1.9.9.2 [55] и провели поиск с помощью ProLuCID [56] в неизбыточной базе данных, содержащей базу данных Ixodidae из Национального центра биотехнологической информации (NCBI, www.ncbi.nlm.nih.gov) (62 246 записей), объединенную с Oryctolagus cuniculus из справочной базы данных Uniprot (www.uniprot.org) (21 148 записей) и обратные последовательности всех записей. Избыточность последовательностей в базе данных была удалена модулем FastaDBXtractor из PatternLab для платформы Proteomics [57]. Поиск проводился с использованием Integrated Proteomics Pipeline – IP2 (Integrated Proteomics Applications, Inc., Сан-Диего, Калифорния, США). Область поиска включала всех кандидатов в пептиды, являющиеся полностью триптическими и полутриптическими. Карбамидометилирование цистеина использовали в качестве статической модификации. Данные были проанализированы с толерантностью к ионам предшественника 50 м.д. и толерантностью к ионам фрагментов 20 м.д.
Достоверность совпадений пептидного спектра (PSM), генерируемых ProLuCID [56], оценивалась с помощью модуля Search Engine Processor (SEPro) от PatternLab для платформы Proteomics [57]. Идентификации были сгруппированы по состоянию заряда и триптическому статусу, в результате чего были выделены четыре отдельные подгруппы.Для каждой группы были использованы ProLuCID XCorr, DeltaCN, DeltaMass, ZScore, количество сопоставленных пиков и вторичные значения ранга для создания байесовской дискриминирующей функции. Пороговая оценка была установлена для принятия уровня ложного обнаружения белков (FDR) в 1% в зависимости от количества ловушек. Эту процедуру выполняли независимо для каждого подмножества данных, в результате чего количество ложных срабатываний не зависело от триптического статуса или состояния заряда. Кроме того, требовалась минимальная длина последовательности из шести остатков на пептид.Результаты были обработаны после обработки, чтобы принять только PSM с ошибкой массы предшественника <10 ppm.
Функциональная аннотация и классификация белков
Поиск по BLASTP в нескольких базах данных был выполнен для аннотирования найденных белков. Для проверки идентичности белков клещей использовались следующие базы данных: неизбыточные (NR), Acari и refseq-беспозвоночные из NCBI, Acari из Uniprot, подмножество GeneOntology (GO) FASTA [58], база данных MEROPS [59] и База данных консервативных доменов NCBI [60], содержащая мотивы COG [61], PFAM [62] и SMART [63].Для проверки белков кроликов использовались следующие базы данных: Oryctolagus cuniculus и базы данных refseq-vertebrates из NCBI, O . cuniculus от Uniprot, подмножество GeneOntology (GO) FASTA [58], база данных консервативных доменов NCBI [60], содержащая мотивы COG [61], PFAM [62] и SMART [63]. Для функциональной классификации белковых последовательностей использовалась программа доктора Хосе М. С. Рибейро, написанная на Visual Basic 6.0 (Microsoft, Редмонд, Вашингтон, США) [34].Функционально аннотированный каталог для каждого набора данных был вручную обработан и введен в электронную таблицу Excel с гиперссылками (таблицы S1 и S2).
Относительное обилие и графическая визуализация
Для определения относительного содержания белков слюны использовали нормированные коэффициенты спектрального содержания (NSAF). Значение NSAF было подтверждено как надежное в методе относительной количественной оценки без меток [64–66]. Использовали средние NSAF из двух или трех повторов. Для определения относительной численности среднее значение NSAF для каждого функционального класса белка или отдельного аннотированного белка выражали в процентах (%) от общего количества NSAF на момент времени.Чтобы визуализировать паттерны относительной экспрессии на тепловой карте, значения% NSAF были нормализованы с использованием статики Z-баллов с использованием формулы, где Z i s – Z-оценка, X – NSAF для каждого белка в момент времени, μ. – это среднее значение по временным точкам, σ – стандартное отклонение по временным точкам. Нормализованные значения NSAF использовались для создания тепловых карт с использованием функции heatmap2 из библиотеки gplots в R [67].
Филогенетический анализ
аминокислотных последовательностей были использованы для построения направляющего дерева филогении с помощью MacVector 12.7.3 (MacVector Inc Cary, Северная Каролина, США). Последовательности белков были выровнены с использованием метода Muscle в MacVector с настройками по умолчанию. Впоследствии дерево было построено с использованием метода Neighbor Joining с нескорректированной («p») настройкой расстояния. Для оценки значений начальной загрузки было установлено 1000 репликаций.
Результаты и обсуждение
Сбор слюны клеща
Мы успешно получили слюну, индуцированную пилокарпином I . scapularis клещей, которые частично кормились кроликами в течение 24, 48, 72, 96 и 120 часов, а также клещей, которые явно налились кровью, но не отделились (BD), и клещей, которые налились кровью и спонтанно отделились (SD).Во время сбора слюны мы наблюдали, что слюна клещей, получавших питание в течение 24 часов, быстро высыхала, образуя хлопьевидные белые кристаллические остатки, и для сбора мы растворили эти хлопья в 2 мкл стерильного физиологического раствора с фосфатным буфером (PBS, pH 7,4). Напротив, капля слюны клещей на последующих этапах кормления была видна в течение нескольких секунд или минут после инъекции пилокарпина.
Белковый состав в
I . scapularis слюна клеща смена каждые 24 ч В таблицеS1 перечислены белки клещей и кроликов, которые были идентифицированы в I . scapularis слюна. Поиск извлеченных тандемных масс-спектров по базе данных белков клещей и кроликов с использованием ProLucid [56] и фильтрация с использованием SEPro [57] позволили получить совпадения с 769 белками клещей и 130 белков кролика соответственно с по крайней мере одним совпадением пептида на белок (таблица S1, обратите внимание разные вкладки). При дальнейшем анализе в модуле BirdsEye View от PatternLab для платформы Proteomics [57] 582 из 769 клещевых белков были определены как аутентичные, поскольку они были обнаружены в двух или всех трех прогонах, в то время как остальные 187 белков были обнаружены только в один из трех прогонов был признан совпадением с низкой достоверностью и в дальнейшем не обсуждался (таблица S1).Из 130 кроличьих белков, обнаруженных в I . scapularis слюна клеща, 83 соответствовали критериям аутентификации. При автоматической аннотации [34] 582 клещевых и 83 кроличьих белка с высокой степенью достоверности, соответственно, классифицируются на 24 (таблица 1) и 18 (таблица 2) функциональных классов белков. В частности, в таблицах 1 и 2 суммированы совокупные количества белков, которые были идентифицированы в каждом функциональном классе, очевидное относительное количество в каждый момент времени и моменты времени, в которых класс не был обнаружен [представлен нулем (0)].
На рис. 1 и 2 представлены снимки относительного обилия белков клеща (рис. 1) и кролика (рис. 2) в I . scapularis слюна каждые 24 часа. На рис.1 видно, что большинство I . scapularis клещевые белки в этом исследовании принадлежат к четырем преобладающим функциональным классам белков, начиная с белков с неизвестной функцией, за которыми следуют ингибиторы протеаз (PI), антимикробные / иммунные белки и гем-связывающие белки. Далее следуют классы белков с низким содержанием, на долю которых приходится 1–6% (цитоскелет, богатый глицином и механизмы модификации белков), при этом оставшиеся обнаруженные классы белков составляют менее 1%.Из четырех основных классов белков относительное количество белков с неизвестной функцией, по-видимому, увеличивается с кормлением, составляя 33–58% от общего белка в период между 24–120 часами, а затем снижается до 13% в слюне полностью накормленных, но не отделившихся клещей (BD ), а также полностью сытые и самопроизвольно отделившиеся. Точно так же гемсвязывающие белки увеличились с ~ 14% через 24 часа до ~ 24% через 96 часов, затем упали до 10% через 120 часов, снова поднялись до 24% в BD и упали до 8% в SD. С другой стороны, количество ИП и антимикробных / связанных с иммунитетом пептидов снижалось при кормлении, а количество первых снижалось с 24.От 7% через 24 часа до 17–8% через 48–120 часов соответственно, но возрастает до 21% в BD и снижается до 7% в SD. Аналогичным образом антимикробные / связанные с иммунитетом белки снизились с 18% через 24 часа до 17–5% через 48–120 часов, а затем несколько повысились до ~ 7% при BD и SD (Таблица 1 и Рис. 1). Известные классы белков включают протеазы и липокалины, количество которых увеличивается с кормлением. Содержание протеазы увеличивается с 0,5% через 24 часа до 3–6% через 48, 96 и 120 часов, за исключением 72 часов, когда содержание было 0,5%, и 5–6% в BD и SD (рис. 1).Аналогичным образом содержание липокалина увеличивается с 0,3% через 24 часа до ~ 6% через 120 часов, что не обнаруживается в BD, но составляет ~ 3% от содержания белка в SD. Также видно из Таблицы 1 и Рис. 1, количество белков, подобных домашнему хозяйству клещей, увеличивается с кормлением.
Рис. 1. Относительная численность классов белков клещей в I . scapularis слюна клеща во время и после кормления.
Общий нормализованный коэффициент спектрального изобилия (NSAF) для каждого класса белков выражается как процент от общего NSAF на момент времени.Предоставляется ключ, в котором перечислены 24 класса белков, идентифицированных в слюне клещей как белки, полученные от клещей.
https://doi.org/10.1371/journal.pntd.0004323.g001
Рис. 2. Относительная численность классов белков хозяина (кролика) в I . scapularis слюна клеща во время и после кормления.
Общий нормализованный коэффициент спектрального изобилия (NSAF) для каждого класса белков выражается как процент от общего NSAF на момент времени. Предоставляется ключ, в котором перечислены 18 классов белков, идентифицированных в слюне клещей как белки, производные от хозяина.
https://doi.org/10.1371/journal.pntd.0004323.g002
На рис. 2 показано относительное содержание кроличьих белков, обнаруженных в I . scapularis слюна. Интересно отметить, что из 18 классов белков в Таблице 2 четыре класса белков: гем / железо, продукты разложения гемоглобина / эритроцитов, антимикробные вещества / связанные с иммунитетом и кератин были обнаружены во всех временных точках. Примечательно, что эти четыре класса белков представляют собой наиболее распространенные белки кроликов в слюне клещей.За исключением 72 часов, когда кроличьи гем / железосвязывающие белки составляли 3%, этот белок был одним из наиболее преобладающих в другие моменты времени, составляя 17–75% от общего содержания кроличьего белка в слюне клещей. Точно так же содержание белков, связанных с гемоглобином / эритроцитами, увеличилось с 7,6% через 24 часа до 50,2% при BD и 39,3% при SD. Связанные с иммунитетом белки кроликов были наиболее распространены через 48, 96 и 120 часов слюны – 30%, 29,6% и 27,6% соответственно (рис. 2). Кератины, обнаруженные во всех временных точках, могут сигнализировать о загрязнении наших образцов при обращении.Еще одно интересное наблюдение на рис. 2, фибриноген, предшественник фибрина, который участвует в образовании сгустка, был обнаружен к концу кормления, 0,25% в 96-часовой слюне возрастает до 3,1% в SD. Может ли это означать, что клещ поглощает фибриноген во время кормления и секретирует его обратно хозяину во время фазы отслоения, чтобы способствовать заживлению ран? Твердые клещи создают рану на коже хозяина, из которой они высасывают кровь, однако эта рана полностью заживает, когда клещи отслаиваются.
Динамика секреции выбранных классов белков в течение
I . scapularis кормление клещейМы заинтересованы в понимании механизмов, которые регулируют питание клещей на ранней стадии, и поэтому последующее обсуждение данных смещено в сторону белков, полученных из клещей, не связанных с домашним хозяйством, которые были обнаружены в слюне после 24/48 часов. Мы обсудили белки, полученные из кроликов, отдельно, но при необходимости подчеркнем сходства и различия. Мы использовали нормализацию Z-статистики значений NSAF (таблица S3) для построения тепловых карт на рисунках 3–5. Эти данные дают представление об относительном количестве специфических белков во время кормления: протеазы (рис. 3A), ингибиторы протеаз (рис. 3B), липокалины / гистамин-связывающие белки клещей / связывающие жирные кислоты белки (рис. 4A), антимикробные / иммунные. (Рис. 4B), гем-связывающие белки (Рис. 4C), антиоксиданты (Рис. 4D), белки с неизвестной функцией (Рис. 5A), богатые глицином белки (Рис. 5B) и белки внеклеточного матрикса (Рис. 5C).
Рис. 3. Динамика секреции протеаз и ингибиторов протеаз в I . scapularis слюна клеща во время кормления.
Нормализованный спектральный коэффициент изобилия (NSAF) для каждого белка в качестве заместителя для относительной численности выражается в процентах от общего количества NSAF на момент времени в пределах каждого класса. Z-баллы были рассчитаны и использованы для создания тепловых карт, как описано в разделе «Материалы и методы». Красный цвет указывает на белки с высоким содержанием, а синий цвет указывает на белки с низким содержанием, количество как увеличивается, так и уменьшается с интенсивностью цвета.Дендрограммы показывают кластеризацию белков (C) в соответствии с паттернами секреции. Ветви помечены, начиная с буквы класса белка. Фиг.3A (металлопротеазы) и фиг.3B (ингибиторы протеаз, B1 = серпины, B2 = ингибиторы протеаз домена TIL, B3 = α-2-макроглобулин и B4 = цистатины) сгруппированы по функциональным классам.
https://doi.org/10.1371/journal.pntd.0004323.g003
Рис. 4. Динамика секреции I . scapularis белки слюны клеща, связанные с противовоспалительными (липокалины), антимикробными, гемосвязывающими и антиоксидантными функциями.
Нормализованный спектральный коэффициент изобилия (NSAF) для каждого белка в качестве заместителя для относительной численности выражается в процентах от общего количества NSAF на момент времени в пределах каждого класса. Z-баллы были рассчитаны и использованы для создания тепловых карт, как описано в разделе «Материалы и методы». Красный цвет указывает на белки с высоким содержанием, а синий цвет указывает на белки с низким содержанием, количество как увеличивается, так и уменьшается с интенсивностью цвета. Дендрограммы показывают кластеризацию белков (C) в соответствии с паттернами секреции.Ветви помечены, начиная с буквы класса белка. Фиг. 4A (липокалины), фиг. 4B (антимикробное средство), фиг. 4C (связывание гема) и фиг. 4D (антиоксиданты) сгруппированы по функциональным классам.
https://doi.org/10.1371/journal.pntd.0004323.g004
Рис. 5. Динамика секреции I . scapularis белки слюны клеща неизвестной функции, богатые глицином и белки, связанные с внеклеточным матриксом.
Нормализованный спектральный коэффициент изобилия (NSAF) для каждого белка в качестве заместителя для относительной численности выражается в процентах от общего количества NSAF на момент времени в пределах каждого класса.Z-баллы были рассчитаны и использованы для создания тепловых карт, как описано в разделе «Материалы и методы». Красный цвет указывает на белки с высоким содержанием, а синий цвет указывает на белки с низким содержанием, количество как увеличивается, так и уменьшается с интенсивностью цвета. Дендрограммы показывают кластеризацию белков (C) в соответствии с паттернами секреции. Ветви помечены, начиная с буквы класса белка. Фиг. 5A (белки слюны клеща с неизвестной функцией), фиг. 5B (богатый глицином) и фиг. 5C (внеклеточный матрикс) сгруппированы по функциональным классам.
https://doi.org/10.1371/journal.pntd.0004323.g005
Большинство протеаз в
I . scapularis слюна металлопротеазМодель I . scapularis Геном кодирует по меньшей мере 233 предположительно активных и 150 предположительно неактивных протеаз, принадлежащих к семействам сериновых, цистеиновых, аспарагиновых, металло- и треониновых протеаз [68]. В этом исследовании мы обнаружили 33 протеазы в четырех кланах: серин- (n = 3), цистеин- (n = 3), аспарагиновые- (n = 1) и металлопротеазы (n = 26) (таблица S2).При поиске по базе данных Merops [59], 26 металлопротеиназ принадлежат семействам M12 (n = 15), M20 (n = 4), M2 (n = 2), M28 (n = 2), M13 (n = 1). , M17 (n = 1) и M49 (n = 1) (таблица S2), в то время как сериновые, цистеиновые и аспарагиновые протеазы классифицируются в семействах S1, C1 и A1 соответственно. Большинство протеаз здесь, вероятно, связано с регуляцией питания клещей: 75% (25/33) были обнаружены между 24–120 часами во время кормления клещами, за исключением семи, которые были идентифицированы только в SD (таблица S2).Около 40% протеаз в составе I . scapularis геном – металлопротеиназы [68]. Независимо от того, обнаружено ли в этом исследовании, что большинство протеаз в I . scapularis слюна клеща является металлопротеазой, отражает протеазный состав в I . scapularis или это физиологическое событие, на данный момент неизвестно.
Статистический анализZ-показателя и визуализация нормализованных значений NSAF 26 металлопротеаз (рис. 3A) показывают, что металлопротеиназы M12 и M2, вероятно, секретировались в большом количестве в период между 24–120 часами во время кормления соответственно, что может указывать на важность этих белков для регулирование первых пяти дней кормления клещей.Остальные металлопротеиназы в семействах M17, 20, 28 и 49, которые были в изобилии в BD и SD (рис. 3A), вряд ли связаны с регулированием событий кормления клещей.
Наблюдение, что I . scapularis , преимущественно секретируемые металлопротеиназами семейства M12 во время кормления, свидетельствует о важности этого класса белков в физиологии питания клещей. Новые свидетельства указывают на то, что это так. Рекомбинантный белок протеазы M12 (AAP22067.1, MCC Fig. 3A) обладает желатиназной и фибрин (оген) олитической активностями [69], что является фактором кормления клещей.В родственном исследовании было выявлено подавление РНКи гомологов AAM
Большинство ингибиторов протеаз в
I . scapularis слюна, вероятно, ингибирует сериновые протеазыПервая линия защиты хозяина от кормления клеща, такая как воспаление, агрегация тромбоцитов, свертывание крови, активация комплемента и клеточный иммунитет, опосредуются протеазами, которые контролируются ингибиторами протеаз (ИП). С этой точки зрения было выдвинуто предположение, что клещи могут вводить ИП в хозяина, чтобы избежать защиты хозяина [18, 74–77].В этом исследовании мы идентифицировали 43 предполагаемых ИП (таблица S1), которые, согласно базе данных Merops, принадлежат к восьми семействам: I2 (ингибиторы сериновой протеазы типа Кунитца, n = 2), I4 (ингибиторы сериновой протеазы, [серпины], n = 11), I8 (ингибиторы сериновой протеазы домена TIL, n = 13), I25 (цистатины, ингибиторы цистеиновой протеазы, n = 4), I31 (тиреопины, ингибиторы цистеиновой протеазы n = 2), I39 (α-2 макроглобулин, A2M, n = 9), I43 (ингибиторы сериновой протеазы типа Kazal, n = 1) и I68 (ингибиторы карбоксипептидазы, TCI, n = 1) были идентифицированы в I . scapularis слюна (таблица S2). Примечательно, что 84% (36/43) ИП были обнаружены в 24- и 48-часовой слюне (таблица S2), что свидетельствует о способности этих белков регулировать ранние стадии питания клещей. Наблюдение здесь, что большинство ИП в этом исследовании, вероятно, являются ингибиторами сериновых протеаз, может сигнализировать о потенциале того, что большинство путей защиты хозяина от кормления клещей, вероятно, опосредованы сериновой протеазой.
Подобно другим классам белков в этом исследовании, относительное количество ИП изменялось каждые 24 часа (рис. 3В).Серпины демонстрируют три профиля секреции: белки SCA в изобилии в первые 48 часов и уменьшаются с кормлением, SCB в изобилии через 96 часов и увеличивается в слюне SD, а количество белков SCC увеличивается с 24 до 120 часов (рис. 3B1). Точно так же ИП домена TIL разделяются на три кластера: те, которые присутствуют в большом количестве в течение первых 48 часов кормления, но уменьшаются при кормлении в TCA, увеличиваются при кормлении между 24–96 часами в TCB, и те, которые преобладают в слюне BD и SD в TCC (рис. 3B2) . На рис. 3B3 альфа-2-макроглобулины разделяются на два кластера: секретируемые в изобилии между 24–120 часами в αCA и 48-SD в αCB.На рис. 3B4 цистатины объединяются в CCA для тех, чья численность увеличивается при кормлении, и CCB для тех, которые секретируются в большом количестве в момент времени 120 часов.
Имеются данные о том, что некоторые ИП, идентифицированные в этом исследовании, регулируют важные функции питания клещей. Например, серпин EEC19556.1 в SCA (рис. 3B1) на 98% идентичен AID54718.1, ингибитору трипсина и тромбина, который также ингибировал свертывание крови и агрегацию тромбоцитов [43]. Аналогично I . ricinus serpin ABI
Липокалины / гистамин-связывающие белки клещей (tHBP) / связывающие жирные кислоты белки (FABP)
Липокалины / HBP и FABPs принадлежат к суперсемейству калицинов гидрофобных лиганд-связывающих внеклеточных белков [87–89]. Семейство белков липокалина, к которому принадлежат HBP, представляет собой большую группу белков, которые связывают и транспортируют небольшие гидрофобные молекулы, а также связаны с множеством функций, включая регуляцию воспаления посредством связывания провоспалительных молекул, таких как гистамин [90–92].Аналогичным образом FABP связывают и транспортируют гидрофобные лиганды, включая длинноцепочечные жирные кислоты, эйкозаноиды, соли желчных кислот и пролифераторы пероксисом [93]. Считается, что липокалины клещей / связывающие гистамин белки участвуют в опосредовании уклонения клещей от защиты хозяина от воспаления за счет секвестрации провоспалительных биогенных аминов, липидов, гистамина, серотонина и простаноидов [94]. Белки, связывающие гистамин (tHBP), представляют собой подмножество липокалинов с двумя карманами для связывания гистамина [91].Из 18 белков в таблице S2 и на рис. 4A 14 аннотированы как tHBP, три – как липокалины и один – как FABP-подобный. Подобно другим белкам, I . scapularis , по-видимому, избирательно вводит tHBP / липокалины в организм хозяина в определенные периоды времени, при этом два tHBP обнаруживаются через 24 часа в LCC, один через 48 часов и три белка каждый через 72 часа и 96 часов в LCD (рис. 4A). Наибольшее количество tHBP / липокалинов было выявлено через 120 часов в LCA (n = 12), половина из которых приходилась исключительно на этот момент времени (таблица S2 и рис. 4A).Примечательно, что два tHBP и по одному липокалину и FABP-подобному, идентифицированные в этом исследовании, были исключительны для SD слюны в LCB, что может предполагать, что эти белки участвуют в событиях в конце кормления клещей.
Ограниченное количество исследований предполагает, что липокалины / HBP / FABP действительно выполняют функции кормления клещей. Три R . appendiculatus Было предсказано, что tHBP подавляют воспаление во время кормления кровью, что подтверждается их способностью вытеснять гистаминовые рецепторы [90].В других исследованиях D . reticulatus tHBP связывает гистамин и серотонин [95], а Ornithodoros . moubata tHBP, называемый мубатином, продемонстрировал ингибирование индуцированной коллагеном агрегации тромбоцитов [96]. В недавнем исследовании липокалины / HBP / FABPs были идентифицированы среди 24–48 ч A . americanum иммуногенные белки слюны клещей [40], что позволяет предположить, что эти белки являются частью белков слюны клещей, которые придают устойчивость к клещам у повторно зараженных животных.Примечательно, что у Radulovic et al., [40], наряду с липокалинами / HBPs, лейкотриен B4-подобная протеаза также была обнаружена среди 24–48 ч A . americanum иммуногенные белки слюны клещей. Интересно отметить, что I . ricinus , tHBP, известный как LIR6-связанный лейкотриен B4 [97], провоспалительный медиатор и мощный хемоаттрактант нейтрофилов.
Я . scapularis антимикробные белки слюны клещаСтиль кормления клеща, заключающийся в разрыве тканей хозяина и всасывании крови из раненого места кормления, подвергает хозяина воздействию микробных инфекций.С этой точки зрения постулировалось, что клещи вводят антимикробные пептиды в место кормления, чтобы предотвратить заражение места кормления [23, 48]. Множественные антимикробные пептиды были охарактеризованы у клещей, большинство из которых являются дефензинами [98–106], микроплюзин / микроплюзиноподобными [107–109] и гебреаин / гебреаинподобными [110]. В этом исследовании семь из 15 антимикробных пептидов в таблице S2 и на рис. 4B являются микроплюзиноподобными, единственный лизоцим, а остальные характеризуются доменами распознавания патогенов (n = 7).На рис. 4B показаны три модели секреции, где белки ACA были в изобилии в течение 24–120 часов, ACB присутствовали только через 48 часов, а белки ACC увеличивались с 48–96 часов, но были в большом количестве в слюне BD и SD. За исключением микроплюзина [107], который, как было показано, останавливает рост Micrococcus luteu s и Cryptococcus neoformans [111], ничего не известно о роли (ах) большинства антимикробных пептидов в этом исследовании. Примечательно, что большинство антимикробных пептидов в этом исследовании, по-видимому, вводят хозяину в течение первых 48 часов кормления (n = 11) (таблица S2 и рис. 4B).Понимание функций некоторых из этих антимикробных пептидов позволит выявить микробы, которые I . scapularis хотят держаться подальше от места кормления.
Гем-связывающие белки
При полном питании твердые клещи, по оценкам, впитывают кровь хозяина, которая более чем в 100 раз превышает их первоначальный вес [112]. Катаболизм этого огромного количества крови производит большое количество железа и гема [113–115]. И железо, и гем необходимы для нормального функционирования клеток [113, 114].Однако, если их оставить незащищенными, и железо, и гем могут вызывать повреждение клеток из-за окислительного стресса [116, 117]. Постулируется, что клещи предотвращают опосредованное железом и гемом повреждение клеток клещей за счет экспрессии связывающих железо и гем белков, которые играют две роли: связываются и распределяются по клеткам для нормальной физиологии, секвестрируют избыток железа или гема и предотвращают повреждение клеток, вызванное окислительным стрессом [115 ].
Одним из наиболее примечательных наблюдений в этом исследовании является то, что хотя гем-связывающие белки представлены ~ 2.Идентифицировано 6% (15/582) белков, что составляет ~ 11–24% от общего содержания белка (Таблица 1 и Рис. 1). Это может свидетельствовать о том, что метаболизм гема потенциально является физиологической функцией клеща, которую нельзя сбрасывать со счетов. Наблюдение за тем, что все 15 гемсвязывающих белков в этом исследовании, вероятно, вводятся хозяину в течение 24–48 часов после того, как клещ начинает кормиться (таблица S2), предполагает, что этот механизм важен с начала кормления клеща. На рис. 4В наблюдаются три модели секреции: содержание ГКА увеличивается в 120-часовом BD-белке, ГХБ в избытке в 48 и 96 часов, а ГЦК в большом количестве в первые 48 часов, но уменьшается при кормлении.Примечательно, что пять гемсвязывающих белков, которые были обнаружены во всех временных точках (таблица S2), объединяются в кластеры в HCA (рис. 4C), за исключением EEC13578.1. Эти белки составляют до 38% от общего количества NSAF в этом классе, что может свидетельствовать об их значении в физиологии питания клещей.
Интересно отметить, что и железо, и гем-связывающие белки также были обнаружены в большом количестве в слюне D . андерсони [51], R . microplus [50] и H . longicornis [52]. Однако в этом исследовании было обнаружено только последнее. Является ли это уникальным для I . scapularis или то, что железосвязывающие белки были введены на уровне ниже определяемого, требует дальнейшего исследования. Опубликованные данные свидетельствуют о том, что клещ может выводить токсины из гема / железа путем секвестрации в пищеварительных клетках (гемосомах) [118, 119] и гемолимфе [120–122]. Данные этого и других исследований [40, 41, 50, 52, 123], которые показывают секрецию гемсвязывающих белков в слюне клещей, предполагают третью возможность удаления гема через слюну клещей.Учитывая, что гем обладает провоспалительными функциями [124], секреция этих белков в слюне клещей может быть связана с секвестрацией гема и, таким образом, позволяет клещам уклоняться от защиты хозяина от воспаления. Секвестрация железа является одним из видов антимикробной защиты млекопитающего-хозяина. Чтобы противостоять защите организма хозяина от секвестрации железа, микробы разработали сложные способы связывания железа из окружающей среды [125–127] и непосредственного поглощения гема, который затем переваривается для высвобождения связанного железа [128]. С этой точки зрения возможно, что секреция гемсвязывающих белков является стратегией клеща, чтобы сделать гем доступным для передаваемых патогенов в месте кормления клеща.Здесь важно отметить, что B . burgdorferi , самый главный I . scapularis , передаваемый агентом TBD человека, может не потребовать железа для колонизации хозяина [129].
Антиоксиданты
Повреждение ткани, вызванное кормлением клеща, такое как разрушение ткани хозяина и последующее высасывание крови из раненой области, приведет к выработке активных форм кислорода (ROS), которые, в свою очередь, повредят ткань хозяина и / или передают возбудителей TBD [130, 131] .Таким образом, ожидается, что клещи будут вводить антиоксиданты в место кормления, как это наблюдалось в этом исследовании. На рис. 4D суммировано относительное содержание 36 предполагаемых антиоксидантных белков, 23 из которых были идентифицированы только в слюне SD (таблица S2) и, вероятно, связаны с событиями к концу кормления клещей. Остальные 13 белков были идентифицированы между 24 часами BD и, вероятно, связаны с регуляцией питания клещей. Тепловая карта на рис. 4D показывает, что различные антиоксиданты были обнаружены в большом количестве в разные моменты времени: ANCA в BD и SD, ANCB через 96 часов, ANCC в SD, ANCD через 48 и 72 часа, ANCE через 24 часа и ANCF в 120 ч.Интересно отметить, что некоторые данные в этом исследовании согласуются с предыдущими наблюдениями. Глутатионпероксидаза (AAK.1) ранее обнаруживалась среди иммуно-доминантных белков в заполненном I . scapularis [132] входит в число 23 антиоксидантов, которые были обнаружены только в слюне SD (таблица S3 и рис. 4D).
Роль антиоксидантов в физиологии клещей остается в основном неизвестной. В недавнем исследовании экспрессия гена тиоредоксинпероксидазы увеличилась в органах B . burgdorferi инфицировано I . ricinus клещей [133], что предполагает участие во взаимодействии клещей и патогенов. Интересно отметить, что в этом исследовании белок тиоредоксинпероксидазы у неинфицированных клещей снижался при кормлении (таблица S3). Было бы интересно определить, могут ли антиоксидантные белки, идентифицированные в этом исследовании, играть роль на интерфейсе клещ-хозяин в приобретении и передаче TBD.
Я . scapularis белки слюны клеща неизвестной функцииБолее 30% последовательностей клещей в общедоступных базах данных имеют неизвестную функцию [30, 32–37, 48, 134–138].В этом исследовании мы идентифицировали 129 белков слюны клещей (TSP) неизвестной функции (таблица S2). Для ясности профили секреции 112 TSP с неизвестной функцией суммированы на фиг. 5A, тогда как остальные 17 белков, богатых глицином, которые, как полагают, участвуют в образовании цемента клещей [139], показаны на фиг. 5B. Интересно отметить, что в таблице S2 93,7% (105/112) TSP были обнаружены в 24–120-часовой слюне, что может указывать на то, что эти белки важны для физиологии питания клещей.Остальные 6,3% (7/112) относятся исключительно к стадиям BD и SD и, вероятно, связаны с событиями ближе к концу кормления. Некоторые белки были обнаружены в один момент времени: 48 (n = 12), 72 (n = 7), 96 (n = 5) и 120 ч (n = 14) слюны (таблица S2). Более половины (n = 62) TSP неизвестной функции были обнаружены в течение первых 48 часов кормления. Это может иметь решающее значение для начала кормления и развития клещей. Паттерны на рис. 5A предполагают, что клещ потенциально может выборочно вводить различные белки своему хозяину каждые 24 часа.Таким образом клещ мог успешно уклоняться от иммунитета хозяина и питаться кровью. Наблюдаются семь кластеров (UCA-UCG) TSP неизвестной функции (рис. 5A). Наиболее примечательно то, что TSP с неизвестной функцией, которые очень распространены через 24 часа (UCG рис. 5A), уменьшаются при кормлении, указывая на то, что эти белки могут служить ключевыми белками в начале процесса питания клещей. Другие паттерны секреции включают белки, которые присутствуют в изобилии через 48, 72, 96 и 120 часов в UCE, UCC, UCF и UCA соответственно, поскольку эти белки могут быть важны для поддержания различных фаз процесса питания клещей.Белки в UCD и UCA могут играть важную роль в конце кормления, например, в заживлении ран и отслоении от хозяина, или служить маркерами завершения кормления клеща.
Как и несколько других жестких тиков, I . scapularis клещи выделяют цемент, чтобы надежно закрепиться на коже хозяина в течение длительного периода кормления клеща [1, 139, 140]. Исследования химического анализа показали, что цемент клещей имеет высокое содержание белков, богатых глицином [139]. На этом основании мы предполагаем, что богатые глицином белки в таблице S2 могут быть связаны с образованием клещевого цемента.Первый слой цементного конуса клеща откладывается в течение 5–30 мин после прикрепления клеща, а второй слой начинает формироваться через 24 ч после прикрепления [139]. Интересно отметить, что большинство (n = 13) белков, богатых глицином, были идентифицированы в большом количестве в 24- и 48-часовой слюне (таблица S3 и рис. 5B). Характер секреции белков, богатых глицином, показанный на рис. 5В, предполагает, что клещ чередует секрецию этих белков во время кормления. В частности, белки в GCA наиболее распространены через 24 часа, GCB через 48 часов, GCC через 96 часов, GCD в BD-SD и GCE в 72–120 часах слюны (рис. 5B).Важность белков, богатых глицином, обнаруживаемых в изобилии к концу кормления, на данный момент неизвестна. Однако есть вероятность, что эти белки представляют собой продукты дегенерированных слюнных желез. Будет интересно определить функцию этих белков к концу кормления.
При филогенетическом анализе 40,2% (45/112) TSP неизвестной функции уникальны тем, что они разделяются индивидуально, за ними следуют 7,1% (8/112), которые группируются попарно, а остальные 52.7% (59/112) разделяются на пять кластеров (C) A-E (рис. 6). Согласно ранее описанной классификации I . scapularis белков [136], кластеры CA, CB и CD соответственно классифицируются как основные хвостовые (группа 1, n = 15) или бесхвостые белки (группа 2, n = 10), антагонисты GPIIb / IIIa (группа 9, n = 7) и семейством 7–9 кДа (группа 7, n = 11). TSPs в кластере CC (n = 7) имеют мотивы белков, подобных связыванию инсулина [141], тогда как белки кластера CE богаты лейцином (n = 9), как выявлено при проверке последовательности.На основе аминокислотных мотивов Ribeiro et. др. [136] классифицировали основные хвостовые и основные бесхвостые белки на типы I-III. Из 25 белков СА 44% (11/25) и 20% (5/25) подходят для основных белков хвоста I и II соответственно, а остальные 36% (9/25) подходят для основных белков без хвоста.
Рис. 6. Филогенетический анализ белка слюны клеща неизвестной функции, идентифицированного в I . scapularis слюна во время и после кормления.
Направляющее филогенетическое дерево белков слюны клещей (TSP) с неизвестными функциональными последовательностями было построено с использованием метода Neighbor Joining с бутстраповскими репликами, установленными на 1000.Число на каждом узле представляет значения начальной загрузки, которые показывают уровень доверия к ветви. Сгруппированы пять основных групп: (CA) основные хвостовые или бесхвостые белки, (CB) антагонист GPIIb / IIIa, (CC) инсулин-подобные белки (CD) белки 7-9 кДа и (CE) белки, богатые лейцином.
https://doi.org/10.1371/journal.pntd.0004323.g006
Интересное наблюдение из наших данных состоит в том, что белки, которые сегрегированы вместе в (Рис. 6), были идентифицированы в разные моменты времени (Рис. 7A – 7E) предполагая, что клещ может выборочно секретировать эти белки во время кормления.Как и на рис. 3–5, мы использовали нормализацию Z-статистики значений NSAF (таблица S3) для построения тепловых карт на рис. 7A – 7E. Основные хвостовые или бесхвостые белки, разделенные на пять кластеров в соответствии с паттернами секреции, начиная с одиночного белка в BCA, который присутствует в большом количестве в SD, за которым следуют белки в BCB, BCC, BCD и BCE, которых соответственно много в 96, 48, 120 и 72. h слюна (рис. 7A). Аналогичным образом на фиг. 7B, белок-антагонист GPIIb / IIIa разделен на три группы: в изобилии с 72 часов (GPCA), через 24 и 48 часов (GPCB) и только через 48 часов (GPCC).На рис. 7C, за исключением одного белка, которого много в слюне SD (ICB), большинство этих белков присутствует в 24–72 часах слюны (ICA). На рис. 7D и 7E белки, богатые лейцином 7–9 кДа, были идентифицированы на различных уровнях на протяжении всего кормления.
Рис. 7. Относительное количество белков слюны клещей (TSP) неизвестной функции во время и после кормления.
Нормализованный коэффициент спектральной распространенности (NSAF) для TSP, которые разделены вместе на рис. 6, подвергали статистическому анализу по Z-баллу и использовали для создания тепловых карт, как описано в разделе материалов и методов «Относительная численность и графическая визуализация».Красный цвет указывает на белки с высоким содержанием, а синий цвет указывает на белки с низким содержанием, количество как увеличивается, так и уменьшается с интенсивностью цвета. Паттерны кластеризации дендрограмм были основаны на сходстве паттернов секреции. A = основные хвостовые белки или белки без хвоста, B = антагонист GPIIb / IIIa, C = белки, связывающие инсулиноподобный фактор роста, D = белки 7-9 кДа и E = белки, богатые лейцином.
https://doi.org/10.1371/journal.pntd.0004323.g007
Предполагаемые GPIIb / IIIa в кластере GPCB (рис. 7B) характеризуются мотивом «RGD» и потенциально могут блокировать агрегацию тромбоцитов, блокируя связывание активированных тромбоцитов с фибриноген [142].В недавнем исследовании пептиды, содержащие мотив «NGR», препятствовали связыванию тромбоцитов в состоянии покоя с фибриногеном [143]. Интересно отметить, что четыре (AAY66799, AAY66507, AAY66621 и AAY66504) основных белков хвостовой слюны имеют этот мотив. Могут ли эти белки функционально блокировать агрегацию тромбоцитов в состоянии покоя, требует проверки. Если они функционируют, они могут сыграть ключевую роль в успешном кормлении клещей, так как в начале кормления клещ столкнется с покоящимися тромбоцитами. Неожиданно ни один из четырех белков с мотивом NGR не был обнаружен в 24-часовой слюне, когда мы ожидаем покоя тромбоцитов в месте питания.Интересно, что за исключением AAY66507.1 (UCE), обнаруженного в 48- и 96-часовой слюне, остальные три были обнаружены в отдельные моменты времени: AAY66799.1 через 72 часа (UCC), AAY66621.1 через 96 часов (UCF) и AAY66504. .1 через 120 ч (UCB).
Я . scapularis белки, подобные внеклеточному матриксу слюны клещаПодобно белкам, богатым глицином, белки внеклеточного матрикса, вероятно, участвуют в образовании цемента клещей и / или функции клеточной адгезии. В этом исследовании мы обнаружили 9 внеклеточных белков (таблица S2), которые включали кутикулу и хитиназоподобные белки.Два паттерна секреции наблюдаются на рис. 5C, где белки ECB были в изобилии в течение 24–96 часов, а белки ECA были в изобилии в слюне BD и SD. Белки этого класса в 120-часовой слюне не обнаружены. Интересно отметить, что как активная, так и неактивная формы хитиназы были идентифицированы в первые 48 часов. Первый выделяется хитин-связывающим доменом перитрофина-А, который участвует в ремоделировании экзоскелета хитинового клеща, особенно ротовой части [144, 145]. Последний очень идентичен A . americanum пищевые стимулы клещей реагируют на кислую хитиназу [134], которая, когда она подавляется РНКи, заставляет клещей свободно прикрепляться к коже хозяина [146]. Выравнивание Blast2seq показало, что два файла I . scapularis неактивные хитиназы (EEC01936.1 и JAB70416.1), идентифицированные в 24- и 48-часовой слюне, соответственно, на 64 и 65% идентичны A . americanum неактивная хитиназа (AIR
.1). Независимо от того, I . scapularis Неактивные хитиназы выполняют аналогичную функцию во время кормления клещей и требуют дальнейшего изучения.
Белки домашнего хозяйства и другие TSP
В дополнение к антиоксидантам, рассмотренным выше, белки, подобные домашним, идентифицированные в этом исследовании, включают белки, связанные с метаболизмом липидов (n = 15), углеводов (n = 20), промежуточных продуктов (n = 1), энергии (n = 45). ), нуклеотиды (n = 14) и аминокислоты (n = 20) (таблица S2). Другие классифицируются как цитоскелет (n = 32), протеасомный аппарат (n = 10), модификация белка (n = 49), синтез белка (n = 24), экспорт белка (n = 10), ядерная регуляция (n = 7). , сигнальная трансдукция и апоптоз (n = 8), аппарат транскрипции (n = 8), а также переносчики и рецепторы (n = 19) (таблица S2).Накопленный NSAF как показатель обилия белка предполагает, что большинство белков, подобных домашнему хозяйству, секретировалось к концу кормления клещей в слюне BD и SD, соответственно (Таблица 1).
Слюнная железа клеща начинает дегенерировать ближе к концу кормления клеща и почти завершается в течение четырех дней после отделения клеща [140, 147]. Учитывая, что большинство генов домашнего хозяйства функционирует внутри клетки, можно утверждать, что высокое содержание этих белков в слюне BD и SD может представлять прогрессивную деградацию SG к концу кормления клеща.Однако недавний иммуно-скрининг библиотек экспрессии фагового дисплея с использованием антител к 24 ч I . scapularis [41] и 24–48 ч A . americanum [40] белки слюны клещей, которые идентифицировали как домашние белки, указывают на то, что секреция некоторых домашних белков начинается задолго до дегенерации слюнных желез клещей, и, таким образом, эти белки, вероятно, играют важную роль (-и) в регуляции питания клещей.
Одна из замечательных адаптаций клещей заключается в том, что, хотя клещи питаются из раненой области на коже хозяина, место кормления полностью заживает, когда клещи завершают питание и отделяются от кожи хозяина.Существует вероятность того, что некоторые из белков, идентифицированных в BD и SD, могут быть связаны с ускорением заживления ран. Интересно отметить, что некоторые цитоскелетные белки, включая актин [148–152], профилин [153, 154], альфа-тубулин [154], кальпонин [155, 156], немышечный миозин [149, 157, 158], тимозин [159], а тропомиозин [160], выявленные в большом количестве в слюне BD и SD, были связаны с различными аспектами заживления ран. Может ли секреция этих белков в большом количестве быть способом клеща помочь хозяину вылечиться?
Белки-хозяева в
I . scapularis слюнаКогда клещи питаются кровью, они поглощают тысячи белков хозяина. Наблюдение в этом исследовании, что I . scapularis секретировал 83 из тысяч белков-хозяев, что позволяет предположить, что клещ обладает механизмом избирательной секреции белков-хозяев в слюне. Подобно динамике секреции белков, полученных из клещей, клещ, по-видимому, избирательно секретирует различные белки кролика в разные моменты времени кормления клещей (таблица S1). Потенциально возможно, что, подобно белкам, полученным из клещей, белки хозяина в слюне клещей выполняют функции, уникальные для различных фаз питания клещей.Белки, идентифицированные по слюне 24/48 часов и в другие моменты времени (иммунитет / антимикробная функция, метаболизм гема / железа, гемоглобин, ядерная регуляция, внеклеточный матрикс и цепь альфа-1 коллагена), вероятно, помогают клещу питаться. С другой стороны, белки, идентифицированные в слюне BD и SD, такие как фибриноген и ингибиторы протеаз, вероятно, связаны с событиями ближе к концу кормления клеща. Например, функционально аннотированные антимикробные пептиды: антимикробный белок CAP18 [161, 162], идентифицированный во всех образцах, кроме 120-часовой слюны, липокалин, связанный с желатиназой нейтрофилов [163, 164], белок гранул нейтрофилов [165–167], белок S100-A12 [ 168], нейтрофильный антибиотик [165] и лизоцим C [169, 170], которые были идентифицированы через 48 часов и слюна другой стадии (таблица S2), могли помочь клещу избавиться от микробов с места кормления клеща.Интересно отметить, что мы идентифицировали антимикробные пептиды клещевого и кроличьего происхождения в одни и те же моменты времени. Наиболее вероятно, что эти противомикробные пептиды нацелены на различные микробы с помощью белков клещевого происхождения, очищающих от микробов клещевого происхождения, тогда как белки, полученные от хозяина, очищают микробы от хозяина. Было показано, что бесклеточный гемоглобин (Hb) обладает антимикробной активностью в результате окислительного шока [171, 172], и, таким образом, существует вероятность того, что Hb, обнаруженный в слюне клещей, может обеспечивать антимикробную функцию [173, 174] в месте кормления клещей.В другом исследовании пептиды, полученные в результате переваривания гемоглобина протеазами клещей, были описаны как антимикробные пептиды [173, 175, 176]. Также возможно, что секреция гемоглобина может быть признаком переваривания пищи с кровью.
Подобно клещам (рис. 1), белки, связанные с метаболизмом гема и железа кролика, были в большом количестве во все временные точки (рис. 2). Заметное отличие состоит в том, что в то время как мы идентифицировали гем-связывающие белки исключительно для клещей, мы идентифицировали большинство железосвязывающих белков для кроликов (n = 6) и один гем-связывающий белок (таблица S2).Может ли это означать, что I . scapularis использует белки хозяина для удаления избытка железа через слюну? Если да, то это могло быть I . scapularis гемосвязывающие белки клеща ответственны за удаление гема, но клещ задействует связывающие железо белки хозяина для удаления избытка железа. За исключением гаптоглобина [177], который был обнаружен в слюне SD, все другие железосвязывающие белки: сывороточный альбумин, гликопротеин, богатый гистидином, лактотрансферрин и серотрансферрин, а также гем-связывающий белок, гемопексин и сывороточный альбумин были идентифицированы в течение 24/48 часов. слюна и другие стадии во время кормления (таблица S2).По нашим данным, I . scapularis , по-видимому, может использовать белки хозяина для удаления избытка железа из организма хозяина, начиная с 24–48 часов. Также интересно отметить, что человеческий сывороточный альбумин подавляет фактор некроза опухоли альфа (TNF), а компонент комплемента C5a запускает респираторный взрыв нейтрофилов [178, 179]. Возможно, повышенная концентрация сывороточного альбумина в месте кормления клещей может выполнять другие функции. Учитывая, что хозяин использует секвестрацию железа в качестве защитного механизма против микробов [180–183], вполне возможно, что манипуляции клеща с хозяином, чтобы перекачивать железо обратно в место питания, могли быть адаптацией к помощи агентам TBD для колонизации хозяина, за исключением организмов, таких как B . burgdorferi , которые не нуждаются в железе для пролиферации [129].
Интересно отметить, что в этом исследовании мы обнаружили фибриноген кролика в слюне BD и SD. Фибриноген является источником фибрина, необходимого для укрепления тромба [184, 185]. Может быть, клещ закачивает фибриноген обратно в хозяина, чтобы помочь изолировать место кормления в конце кормления клеща? Учитывая, что в коже экспрессируется большое количество кератинов [186, 187], существует вероятность того, что кератиновые белки идентифицированы в I . scapularis слюна клеща могла быть связана с обработкой образцов или загрязнением кожи кролика. Важно отметить, что все типы кератина, которые были идентифицированы в I . scapularis слюна клеща в этом исследовании связана с разными слоями кожи [188, 189], и, таким образом, есть большая вероятность, что мы обнаружили остатки на ротовом аппарате клеща. Однако восемь белков кератина, идентифицированные в этом исследовании, составляют менее четверти из 27 кератинов кожи [189]. Вводит ли клещ избирательно кератины и с какой целью – интересный вопрос для будущих исследований.
Выводы и перспективы на будущее
Уникальным вкладом этого исследования является то, что мы впервые попытались идентифицировать белки, полученные от клещей и хозяев, которые обнаружены в I . scapularis слюна клеща каждые 24 часа в течение первых пяти дней кормления, а также ближе к концу кормления. В этом исследовании представлены идентификаторы I . scapularis белки слюны клещей, связанные с регуляцией: (i) ранних событий кормления клещей, таких как прикрепление клещей к коже хозяина и создания очагов кормления, которые предшествуют передаче возбудителей TBD от клещей, (ii) фазы медленного кормления, когда большинство возбудителей TBD являются передается, и клещ готовится к фазе быстрого кормления, и (iii) фазе быстрого кормления, когда клещ питается до насыщения и отделяется от хозяина.Влияние этих данных на будущие углубленные исследования физиологии питания клещей огромно. Например, передача большинства возбудителей TBD происходит по крайней мере через 36–48 часов после прикрепления клеща [190–194]. Что произойдет, если мы проведем иммунизацию против белков слюны клещей в течение 24–48 часов, остановится ли передача возбудителя TBD? С другой стороны, мы идентифицировали белки, которые, по-видимому, секретировались во все моменты времени. В будущих исследованиях было бы интересно определить, регулируют ли эти белки «обязательные» пути? Было бы интересно проверить важность таких белков, используя подход подавления РНКи.Некоторые белки были обнаружены в одном, двух или трех временных точках, могут ли они регулировать функции, уникальные для этого периода кормления клещей?
Интересная повторяющаяся закономерность, наблюдаемая в этих данных, заключается в том, что некоторые функционально похожие, но антигенно уникальные белки были идентифицированы в разные моменты времени кормления. Мы предполагаем, что это может быть стратегией клеща для защиты основных путей от атаки иммунного ответа. Например, иммунный ответ хозяина против 24-часовых белков не повлияет на функции функционально подобных, но антигенно уникальных белков в более поздние моменты времени кормления.По сути, иммунная защита хозяина от кормления клещами будет время от времени перезапускаться и в конце концов не будет эффективной. Таким образом, основные физиологические функции кормления клещей будут продолжаться непрерывно. Может ли этот механизм быть клещевым эквивалентом антигенной вариации, используемой паразитами, такими как трипаносомы, для уклонения от иммунитета хозяина [195–197]? Что произойдет, если мы нацелимся как на кластер функционально похожих, но уникальных в антигенном отношении белков, которые вводятся хозяину в разные моменты времени?
Мы хотели бы предупредить читателя об ограничениях, присущих данному исследованию.Во-первых, в подходах ЖХ-МС / МС существует вероятность того, что преобладающие белки будут маскировать открытие слабо экспрессируемых, но важных белков, и, следовательно, список I . scapularis белки слюны клеща, представленные здесь, не могут быть исчерпывающими. Во-вторых, мы секвенировали белки в слюне, которые были собраны при стимуляции пилокарпином, и еще предстоит выяснить, все ли обнаруженные белки секретируются в физиологических условиях. Однако нас обнадеживают наши выводы о том, что 13% (76/582) от I . scapularis белки слюны клещей в этом исследовании были описаны в других исследованиях протеомов слюны клещей и иммунотранскриптомных исследованиях (таблица S4). Из 76 белков 12 и 13 белков были обнаружены среди иммуногенных белков слюны клещей, которые связывали антитела с 24 ч I . scapularis [41] и 24–48 ч A . americanum белки слюны клеща [40] соответственно. Кроме того, один белок был идентифицирован в I . scapularis нимфы в качестве иммуногенного белка, который связывается с сывороткой человека от укусов клещей [198].Остальные белки были обнаружены в протеомах слюны R . microplus (n = 28, [50]), H . longicornis (n = 22, [52]), D . andersoni (n = 2, [51]), O . moubata (n = 5, [53], секвенирование I . scapularis слюны клеща деградацией по Эдману (n = 4) [48]), секреты других были подтверждены в исследованиях вестерн-блоттинга [42, 43, 199–204].
Я . scapularis Белки в таблице S4 могут представлять собой высококонсервативные белки слюны клеща, которые регулируют важные функции, которые в случае нарушения могут повлиять на клеща.Эти белки могут быть приоритетными кандидатами в будущих исследованиях. Мы хотели бы отметить, что некоторые из белковых последовательностей в этом исследовании взяты из I . ricinus и другие виды клещей. Большинство этих белковых последовательностей имеют гомологи в I . scapularis , которые были исключены как избыточные, когда мы свернули локальную базу данных. Я .