
Статья 284 тк рф с комментариями 2019: Статья 284 ТК РФ с комментариями
Статья 284 ТК РФ с комментариями
Полный текст ст. 284 ТК РФ с комментариями. Новая действующая редакция с дополнениями на 2021 год. Консультации юристов по статье 284 ТК РФ.
Продолжительность рабочего времени при работе по совместительству не должна превышать четырех часов в день. В дни, когда по основному месту работы работник свободен от исполнения трудовых обязанностей, он может работать по совместительству полный рабочий день (смену). В течение одного месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников.
Ограничения продолжительности рабочего времени при работе по совместительству, установленные частью первой настоящей статьи, не применяются в случаях, когда по основному месту работы работник приостановил работу в соответствии с частью второй статьи 142 настоящего Кодекса или отстранен от работы в соответствии с частью второй или четвертой статьи 73 настоящего Кодекса.
Комментарий к статье 284 ТК РФ
Частью 1 комментируемой статьи определена общая допустимая продолжительность рабочего времени по работе по совместительству, которая составляет не более 4 часов в день. Такая продолжительность обусловлена предоставлением работнику права на отдых и его обеспечением.
В то же время ч.1 комментируемой статьи допускает возможность исключения из данного правила, касающегося тех дней, когда работник свободен от исполнения трудовых обязанностей по основному месту работы. В этом случае он может работать по совместительству полный рабочий день (смену). Безусловно, такая возможность зависит от усмотрения работника, а также его способностей и физических возможностей осуществления работы с учетом такой продолжительности.
Помимо этого, согласно ч.1 комментируемой статьи продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников в течение одного месяца (другого учетного периода).
В ч.2 комментируемой статьи содержится исключение из общего правила по ограничению продолжительности рабочего времени при работе по совместительству. Исключение состоит в том, что нормы об ограничении продолжительности рабочего времени не распространяются на те случаи, когда работник по основному месту работы приостановил работу либо отстранен от работы. В таких случаях у работника появляется высвобожденное время для того, чтобы осуществлять работу по совместительству.
Консультации и комментарии юристов по ст 284 ТК РФ
Если у вас остались вопросы по статье 284 ТК РФ и вы хотите быть уверены в актуальности представленной информации, вы можете проконсультироваться у юристов нашего сайта.
Задать вопрос можно по телефону или на сайте. Первичные консультации проводятся бесплатно с 9:00 до 21:00 ежедневно по Московскому времени. Вопросы, полученные с 21:00 до 9:00, будут обработаны на следующий день.
Комментарий к статье 284 ТК РФ. 2016-2019г. ЮрИнспекция
Устраиваетесь на основную работу, сдаёте в отдел кадров все полагающиеся документы вместе с трудовой книжкой. Далее по желанию можете устроиться ещё к примеру на 2-3 работы. Это будет считаться работой по совместительству. Трудовую сдавать туда не требуется, необходимо всего лишь написать заявление и подтвердить свою квалификацию (приложить копию диплома). Запись о том, что Вы работаете по совместительству могут внести в вашу трудовую книжку с вашего желания по основному месту работы. Для этого требуется справка с места работы по совместительству о том, что вы действительно там работаете. Вашу работу по совместительству регулирует глава 44 ТК РФ: Глава 44. ОСОБЕННОСТИ РЕГУЛИРОВАНИЯ ТРУДА ЛИЦ, РАБОТАЮЩИХ ПО СОВМЕСТИТЕЛЬСТВУ Статья 282. Общие положения о работе по совместительству Совместительство – выполнение работником другой регулярной оплачиваемой работы на условиях трудового договора в свободное от основной работы время. Заключение трудовых договоров о работе по совместительству допускается с неограниченным числом работодателей, если иное не предусмотрено федеральным законом. Работа по совместительству
может выполняться работником как по месту его основной работы, так и у других работодателей. В трудовом договоре обязательно указание на то, что работа является совместительством. Не допускается работа по совместительству лиц в возрасте до восемнадцати лет, на тяжелых работах, работах с вредными и (или) опасными условиями труда, если основная работа связана с такими же условиями, а также в других случаях, предусмотренных настоящим Кодексом и иными федеральными законами. Особенности регулирования работы по совместительству для отдельных категорий работников (педагогических, медицинских и фармацевтических работников, работников культуры) помимо особенностей, установленных настоящим Кодексом и иными федеральными законами, могут устанавливаться в порядке, определяемом Правительством Российской Федерации, с учетом мнения Российской трехсторонней комиссии по регулированию социально-трудовых отношений.
Заключение трудовых договоров о работе по совместительству допускается с неограниченным числом работодателей, если иное не предусмотрено федеральным законом. Работа по совместительству
может выполняться работником как по месту его основной работы, так и у других работодателей. В трудовом договоре обязательно указание на то, что работа является совместительством. Не допускается работа по совместительству лиц в возрасте до восемнадцати лет, на тяжелых работах, работах с вредными и (или) опасными условиями труда, если основная работа связана с такими же условиями, а также в других случаях, предусмотренных настоящим Кодексом и иными федеральными законами. Особенности регулирования работы по совместительству для отдельных категорий работников (педагогических, медицинских и фармацевтических работников, работников культуры) помимо особенностей, установленных настоящим Кодексом и иными федеральными законами, могут устанавливаться в порядке, определяемом Правительством Российской Федерации, с учетом мнения Российской трехсторонней комиссии по регулированию социально-трудовых отношений.
 В течение одного месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников. Ограничения продолжительности рабочего времени при работе по совместительству, установленные частью первой настоящей статьи, не применяются в случаях, когда по основному месту работы работник приостановил работу в соответствии с частью второй статьи 142 настоящего Кодекса или отстранен от работы в соответствии с частями второй или четвертой статьи 73 настоящего Кодекса. Статья 285. Оплата труда
лиц, работающих по совместительству Оплата труда
лиц, работающих по совместительству, производится пропорционально отработанному времени, в зависимости от выработки либо на других условиях, определенных трудовым договором.
В течение одного месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников. Ограничения продолжительности рабочего времени при работе по совместительству, установленные частью первой настоящей статьи, не применяются в случаях, когда по основному месту работы работник приостановил работу в соответствии с частью второй статьи 142 настоящего Кодекса или отстранен от работы в соответствии с частями второй или четвертой статьи 73 настоящего Кодекса. Статья 285. Оплата труда
лиц, работающих по совместительству Оплата труда
лиц, работающих по совместительству, производится пропорционально отработанному времени, в зависимости от выработки либо на других условиях, определенных трудовым договором.Налоговый Кодекс РФ последняя редакция — НК РФ 2020-2019 часть 1, часть 2
Глава 34 НК РФ Страховые взносы- Статья 419 Плательщики страховых взносов
- Статья 420 Объект обложения страховыми взносами
- Статья 421 База для исчисления страховых взносов для плательщиков, производящих выплаты и иные вознаграждения физическим лицам
- Статья 422 Суммы, не подлежащие обложению страховыми взносами
- Статья 423 Расчетный период.
 Отчетный период
Отчетный период - Статья 424 Определение даты осуществления выплат и иных вознаграждений
- Статья 425 Тарифы страховых взносов
- Статья 426 Тарифы страховых взносов в 2017 – 2019 годах
- Статья 427 Пониженные тарифы страховых взносов
- Статья 428 Дополнительные тарифы страховых взносов для отдельных категорий плательщиков
- Статья 429 Тарифы страховых взносов для отдельных категорий плательщиков на дополнительное социальное обеспечение членов летных экипажей воздушных судов гражданской авиации, а также отдельных категорий работников организаций угольной промышленности
- Статья 430 Размер страховых взносов, уплачиваемых плательщиками, не производящими выплат и иных вознаграждений физическим лицам
- Статья 431 Порядок исчисления и уплаты страховых взносов, уплачиваемых плательщиками, производящими выплаты и иные вознаграждения физическим лицам, и порядок возмещения суммы страховых взносов на обязательное социальное страхование на случай временной нетрудоспособности и в связи с материнством
- Статья 432 Порядок исчисления и уплаты страховых взносов, уплачиваемых плательщиками, не производящими выплат и иных вознаграждений физическим лицам
 Отчетный период
Отчетный периодБазовые положения НК РФ в редакции 2020 года
 3 НК РФ. Здесь же указаны последние изменения базовых положений в редакции 2020 года.
3 НК РФ. Здесь же указаны последние изменения базовых положений в редакции 2020 года.- На каждое лицо налагается обязанность по уплате установленных Кодексом налогов и сборов. В основе этого положения равенство и всеобщность обложения налогами. Но при этом в расчет берется физическая возможность плательщика уплачивать налог.
- Исключается дискриминационный характер сборов. Также не допускается различное применение налогов, исходя из критериев расы, социального положения, национальности или религии.
- Для каждого вида налога потребуется экономическое основание. Нельзя применять их в произвольном порядке. Исключаются и сборы, не позволяющие реализовать права, закрепленные в конституции.
- Запрещена установка взысканий, идущих вразрез с единым экономическим пространством страны. Налоги не должны каким-либо образом ограничивать беспрепятственное перемещение товаров, финансов или услуг по территории РФ.
- Не допускается наложение обязанности уплаты сборов и прочих платежей, имеющих признаки налогов, но не предусмотренные в НК РФ.
- Все элементы обложения налогом определяются в процессе установления налогов. Законодательство формулируют так, чтобы каждый налогоплательщик понимал, какие платежи, в каком объеме и в какие сроки он должен вносить в казну государства.
- Если существуют противоречия, сомнения относительно порядка взыскания или отсутствие ясности в законодательных актах, они должны толковаться в пользу плательщика налогов или иных взносов.
- Наиболее актуальная редакция 2020 года отражает цивилизованный подход к обложению налогами. Она сформировалась в процессе правовой эволюции страны. Согласно ей взыскание налогов обосновано с точки зрения экономики.

Закреплены специализированные принципы данной отрасли права. В первую очередь они коснулись равенства, справедливости, соразмерности и законности в ходе взыскания средств. Требование справедливости находит отражение в каждом принципе НК РФ. Но главное, что следует отметить в данной редакции – это требование толкования неопределенности в пользу налогоплательщика.
Как складывались налоговые отношения на исторической карте РФ?
Вместе с созданием нового государства (Российской Федерации) в 1991 году началось формирование и налоговой системы страны. Основы были заложены в октябре этого года, а закончилось создание системы в январе следующего года. В этот период установили положения, касающиеся всех основных налогов.
С того момента, как Кодекс был сформирован, его статьи и положения регулярно меняли и исправляли. Это создавало некоторую беспорядочность в налоговых отношениях. Корректировки утверждали в середине года, многие правки имели обратное действие, а ответственности за некорректно поданную финансовую отчетность не было. Отсюда и хронический дефицит бюджета в период 1992-98 гг. Граждане в этот же срок теряли деньги на вложениях в кредитные общества или финансовые пирамиды.
Когда общие положения, наконец, утвердили, появился и полноценный Налоговый Кодекс. Случилось это в 1998 году. Второй раздел Кодекса частично узаконили двумя годами позднее. После изменения пятой статьи в 2008 году, стало понятно, что начинается эра цивилизованных отношений государства и налогоплательщика. Такие правки 2008 года позволили законам, облегчающим положение плательщиков налогов, вступать в силу в момент их опубликования в СМИ. Устранение задержек и стало ключевым моментом поправок.
После изменения пятой статьи в 2008 году, стало понятно, что начинается эра цивилизованных отношений государства и налогоплательщика. Такие правки 2008 года позволили законам, облегчающим положение плательщиков налогов, вступать в силу в момент их опубликования в СМИ. Устранение задержек и стало ключевым моментом поправок.
Также в новой редакции добавили правило, отменяющее обратную силу для актов, ужесточающих правила налогообложения. Положения, делающие оплату сборов проще, а ответственность мягче, согласно этой редакции, имели обратную силу. Так заложили основу для нового Кодекса, где государственные институты лояльнее относились к объектам обложения налогами.
Какие функции содержит в себе НК РФ?
В последней редакции данный нормативный акт выполняет определенный функционал:
- этот документ регулирует создание бюджетов федерального, местного и регионального уровня;
- определяет правила взаимодействия сторон отношений в данной правовой сфере;
- регулирует пополнение бюджета за счет нерезидентов и организаций, зарегистрированных за пределами страны;
- реализует в реальности принцип справедливости обложения налогами, предусматривающий учет доходов и финансового положения плательщика;
- регулирует фискальные операции в части ввоза и вывоза продукции;
- контролирует распределение и обложение налогом доходов.

Главное доказательство эффективности законов в данной отрасли – это своевременность и объем поступлений в бюджеты различных уровней (регион, муниципалитет, государство). Сейчас соблюдение положений такого базового документа, как НК РФ, осуществляет ведомство, подчиненное Минфину страны – ФНС. Реализация перечисленных выше функций также лежит на данной Службе.
Комментарии
Ст. 284.3-1. Налоговый кодекс с комментариями в действующей редакции
1. Налогоплательщик – участник регионального инвестиционного проекта, указанный в подпункте 2 пункта 1 статьи 25.9 настоящего Кодекса (далее в настоящей статье также – участник), при условии, что доходы от реализации товаров, произведенных в результате реализации регионального инвестиционного проекта, составляют не менее 90 процентов всех доходов, учитываемых при определении налоговой базы по налогу в соответствии с настоящей главой, без учета доходов в виде положительных курсовых разниц, предусмотренных пунктом 11 части второй статьи 250 настоящего Кодекса, вправе применять к налоговой базе налоговые ставки в размерах и порядке, которые предусмотрены настоящей статьей. 2. Если иное не предусмотрено настоящей статьей, налоговая ставка, предусмотренная пунктом 1.5-1 статьи 284 настоящего Кодекса, применяется участниками в течение десяти налоговых периодов начиная с налогового периода, в котором впервые одновременно соблюдены следующие условия:
2. Если иное не предусмотрено настоящей статьей, налоговая ставка, предусмотренная пунктом 1.5-1 статьи 284 настоящего Кодекса, применяется участниками в течение десяти налоговых периодов начиная с налогового периода, в котором впервые одновременно соблюдены следующие условия:1) в соответствии с данными налогового учета признана прибыль от реализации товаров, произведенных в результате реализации регионального инвестиционного проекта;
2) налогоплательщиком – участником регионального инвестиционного проекта выполнено требование к минимальному объему капитальных вложений, установленное подпунктом 4.1 пункта 1 статьи 25.8 настоящего Кодекса;3) налогоплательщик – участник регионального инвестиционного проекта обратился в налоговый орган с заявлением о применении налоговой льготы, указанным в пункте 1 статьи 25.12-1 настоящего Кодекса.3. Налоговая ставка налога, подлежащего зачислению в бюджеты субъектов Российской Федерации, может быть установлена в размере от 0 до 10 процентов в течение пяти налоговых периодов начиная с налогового периода, в котором в соответствии с пунктом 2 настоящей статьи начинается применение налоговой ставки, предусмотренной пунктом 1. 5-1 статьи 284 настоящего Кодекса, и не может быть менее 10 процентов в течение следующих пяти налоговых периодов.4. Участники региональных инвестиционных проектов, удовлетворяющих требованиям, установленным абзацем вторым подпункта 4.1 пункта 1 статьи 25.8 настоящего Кодекса, утрачивают право на применение налоговых ставок в размерах и порядке, которые предусмотрены настоящей статьей, начиная с 1 января 2029 года.5. Участники региональных инвестиционных проектов, удовлетворяющих требованиям, установленным абзацем третьим подпункта 4.1 пункта 1 статьи 25.8 настоящего Кодекса, утрачивают право на применение налоговых ставок в размерах и порядке, которые предусмотрены настоящей статьей, начиная с 1 января 2031 года.
5-1 статьи 284 настоящего Кодекса, и не может быть менее 10 процентов в течение следующих пяти налоговых периодов.4. Участники региональных инвестиционных проектов, удовлетворяющих требованиям, установленным абзацем вторым подпункта 4.1 пункта 1 статьи 25.8 настоящего Кодекса, утрачивают право на применение налоговых ставок в размерах и порядке, которые предусмотрены настоящей статьей, начиная с 1 января 2029 года.5. Участники региональных инвестиционных проектов, удовлетворяющих требованиям, установленным абзацем третьим подпункта 4.1 пункта 1 статьи 25.8 настоящего Кодекса, утрачивают право на применение налоговых ставок в размерах и порядке, которые предусмотрены настоящей статьей, начиная с 1 января 2031 года.ст. 284 Трудового Кодекса РФ в текущей редакции и комментарии к ней
Продолжительность рабочего времени при работе по совместительству не должна превышать четырех часов в день. В дни, когда по основному месту работы работник свободен от исполнения трудовых обязанностей, он может работать по совместительству полный рабочий день (смену). В течение одного месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников.
В течение одного месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников.
Ограничения продолжительности рабочего времени при работе по совместительству, установленные частью первой настоящей статьи, не применяются в случаях, когда по основному месту работы работник приостановил работу в соответствии с частью второй статьи 142 настоящего Кодекса или отстранен от работы в соответствии с частью второй или четвертой статьи 73 настоящего Кодекса.
Комментарий к статье 284 ТК РФ
1. В соответствии с ч. 1 комментируемой статьи продолжительность рабочего времени при работе по совместительству не должна превышать четыре часа в день.
В те дни, когда по основному месту работы работник свободен от исполнения трудовых обязанностей, он может работать по совместительству полный рабочий день (смену). Установив такое правило, закон вместе с тем предусмотрел некоторые гарантии, обеспечивающие охрану труда работников. В частности, законом предусмотрено, что в течение месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половину месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников.
Установив такое правило, закон вместе с тем предусмотрел некоторые гарантии, обеспечивающие охрану труда работников. В частности, законом предусмотрено, что в течение месяца (другого учетного периода) продолжительность рабочего времени при работе по совместительству не должна превышать половину месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), установленной для соответствующей категории работников.
Исключение из этого правила в соответствии со ст. 350 ТК установлено в отношении медицинских работников организаций здравоохранения, проживающих и работающих в сельской местности и в поселках городского типа. Согласно Постановлению Правительства РФ от 12.11.2002 N 813 продолжительность их работы по совместительству не должна превышать восемь часов в день и 39 часов в неделю (см. коммент. к ст. 350).
2. Согласно ч. 2 ст. 284 установленные ч. 1 этой статьи ограничения продолжительности рабочего времени при работе по совместительству, т.е. не более четырех часов в день и не более половины месячной нормы рабочего времени (нормы рабочего времени за другой учетный период), не применяются в случаях, когда по основному месту работы работник приостановил свою трудовую деятельность в связи с задержкой выплаты заработной платы на срок более 15 дней (ч. 2 ст. 142 ТК) или отстранен от работы на определенный срок в соответствии с медицинским заключением (ч. ч. 2, 4 ст. 73 ТК) (см. коммент. к указанным статьям).
2 ст. 142 ТК) или отстранен от работы на определенный срок в соответствии с медицинским заключением (ч. ч. 2, 4 ст. 73 ТК) (см. коммент. к указанным статьям).
Другой комментарий к статье 284 ТК РФ
§ 1. В наименование ст. 284 внесено редакционное изменение: слова “на условиях совместительства” заменены словами “при работе по совместительству”.
Существенные коррективы внесены в текст этой статьи. Прежний лаконичный текст заменен более подробной регламентацией продолжительности рабочего времени совместителей.
§ 2. Продолжительность рабочего времени – одно из основных условий труда. Поскольку совместительство дополняет основную работу и может осуществляется только в нерабочее время, свободное от обязанностей по основному трудовому договору, продолжительность времени работы по совместительству в интересах охраны здоровья работника ограничивается законом.
§ 3. Статья 284 в новой редакции содержит не только ограничения продолжительности рабочего времени по совмещаемой работе, но и возможность установления для совместителей полной продолжительности рабочего времени при определенных этой же статьей обстоятельствах и условиях.
Статьей 284 установлены следующие ограничения:
1) продолжительность рабочего времени при работе по совместительству не должна превышать четырех часов в день;
2) в течение одного месяца (другого учетного периода) продолжительность работы по совместительству не должна превышать половины месячной нормы рабочего времени (нормы рабочего времени за учетный период), установленной для соответствующей категории работников.
Работа совместителя на условиях полного рабочего времени возможна:
1) в дни, когда по основному месту работы работник свободен от исполнения трудовых обязанностей;
2) когда по основному месту работы работник приостановил работу в соответствии с ч. 2 ст. 142 ТК;
3) когда работник отстранен от работы в соответствии с ч. 2 или 4 ст. 73 ТК.
§ 4. Продолжительность работы, как и иные условия труда совместителя в указанных в ст. 284 пределах, определяется трудовым договором, заключенным для работы по совместительству.
Чувствительность процесса промерзания активного слоя к снежному покрову в Арктической Аляске
Arcioni, M. , Bensi, P., Fehringer, M., Fois, F., Heliere, F., Lin, C.-C., and
Сципал, К .: Миссия по биомассе, статус спутниковой системы, 2014 IEEE
Симпозиум по геонаукам и дистанционному зондированию, Квебек, Квебек,
1413–1416, https://doi.org/10.1109/IGARSS.2014.6946700, 2014.
, Bensi, P., Fehringer, M., Fois, F., Heliere, F., Lin, C.-C., and
Сципал, К .: Миссия по биомассе, статус спутниковой системы, 2014 IEEE
Симпозиум по геонаукам и дистанционному зондированию, Квебек, Квебек,
1413–1416, https://doi.org/10.1109/IGARSS.2014.6946700, 2014.
Армстронг Р., Бродзик М. Дж., Ноулз К. и Савойя, М .: Global Ежемесячная климатология эквивалента снега и воды EASE-Grid, версия 1, указать использованное подмножество, Боулдер, Колорадо, США, Национальный центр данных по снегу и льду НАСА, Центр распределенного активного архива, https: // doi.org / 10.5067 / KJVERY3MIBPS, 2005.
Банин А. и Андерсон Д. М .: Влияние изменений концентрации соли во время Замораживание незамерзшей воды пористых материалов, водные ресурсы. Res., 10, 124–128, https://doi.org/10.1029/WR010i001p00124, 1974.
Бискаборн, Б.К., Ланкман, Ж.-П., Лантуит, Х., Элгер, К., Стрелецкий, Д.А. , Кабель, WL,
и Романовский, В. Э .: Новая база данных Глобальной наземной сети для
Вечная мерзлота (ГТН-П), Earth Syst.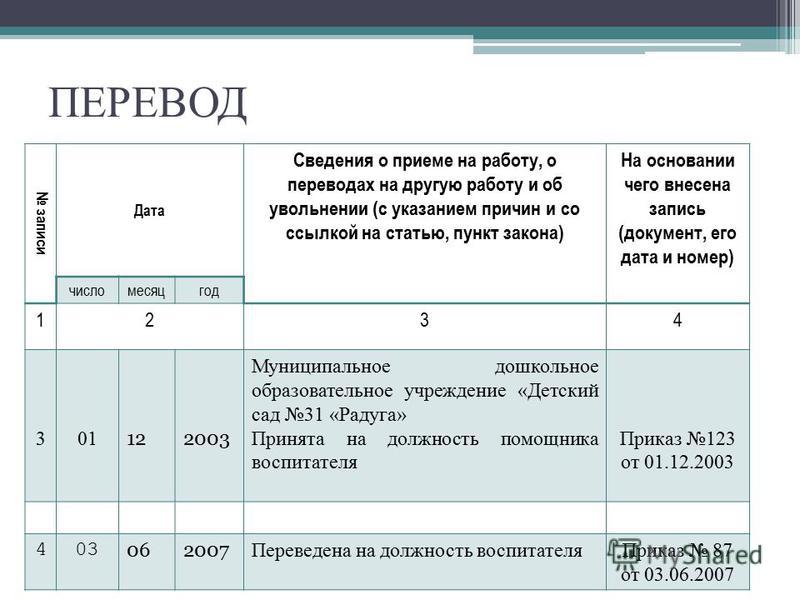 Sci. Данные, 7, 245–259, https: // doi.org / 10.5194 / essd-7-245-2015, 2015.
Sci. Данные, 7, 245–259, https: // doi.org / 10.5194 / essd-7-245-2015, 2015.
Браун Дж., Хинкель К. М. и Нельсон Ф. Э .: Приполярный активный слой. программа мониторинга (CALM): планы исследований и первоначальные результаты, Polar Geography, 24, 166–258, 2000.
Браун, Р., Дерксен, К., и Ван, Л.: Анализ нескольких наборов данных изменчивость и изменение площади весеннего снежного покрова Арктики, 1967–2008 гг., J. Geophys. Res., 115, D16111, https://doi.org/10.1029/2010JD013975, 2010.
Браун, Р. Д. и Дерксен, К. Степень снежного покрова в Евразии в октябре увеличение ?, Environ.Res. Lett., 8, 024006, https://doi.org/10.1088/1748-9326/8/2/024006, 2013.
Берк, Э. Дж., Данкерс, Р., Джонс, К. Д., и Уилтшир, А. Дж .: А ретроспективный анализ вечной мерзлоты в Арктике с использованием поверхности суши JULES модель, Клим. Dynam., 41, 1025–1038, г. https://doi.org/10.1007/s00382-012-1648-x, 2013.
Calonne, N., Flin, F., Morin, S., Lesaffre, B., du Roscoat, S. R. и
Гейндро, Ч .: Численные и экспериментальные исследования эффективных
теплопроводность снега, Геофиз.Res. Lett., 38, L23501,
https://doi.org/10.1029/2011GL049234, 2011.
R. и
Гейндро, Ч .: Численные и экспериментальные исследования эффективных
теплопроводность снега, Геофиз.Res. Lett., 38, L23501,
https://doi.org/10.1029/2011GL049234, 2011.
Chen, R.H., Tabatabaeenejad, A., and Moghaddam, M: P-Band Radar Retrieval свойств активного слоя вечной мерзлоты: подход и проверка временных рядов с наблюдениями на месте IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Валенсия, 6777–6779, https://doi.org/10.1109/IGARSS.2018.8518179, 2018.
Чен, Р. Х., Табатабаинеджад, А., Могхаддам, М .: Восстановление вечной мерзлоты Свойства активного слоя с использованием радиолокационных наблюдений в P-диапазоне временного ряда, IEEE Т.Geosci. Remote, принято, 2019.
Commane, R., Lindaas, J., Benmergui, J., Luus, K. A., Chang, R. Y.-W., Daube, B.C.,
Ойскирхен, Э.С., Хендерсон, Дж. М., Карион, А., Миллер, Дж. Б., Миллер, С. М.,
Паразоо, Н.С., Рандерсон, Дж. Т., Суини, К., Танс, П., Тонинг, К., Веравербек, С.,
Миллер К. Э. и Вофси С. С. Источники углекислого газа на Аляске, вызванные увеличением
раннее зимнее дыхание из арктической тундры, P. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ,
https://doi.org/10.1073/pnas.1618567114, 2017 г.
Источники углекислого газа на Аляске, вызванные увеличением
раннее зимнее дыхание из арктической тундры, P. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ,
https://doi.org/10.1073/pnas.1618567114, 2017 г.
Димс, Дж. С., Пейнтер, Т. Х., и Финнеган, Д. К. Лидарные измерения снега глубина: обзор, J. Glaciol., 59, 467–479, https://doi.org/10.3189/2013JoG12J154, 2013.
Де Ланнуа, Г. Дж. М., Костер, Р. Д., Райхле, Р. Х., Маханама, С. П. П., и Лю, К .: Обновленная обработка текстуры почвы и связанной с ней гидравлической системы. свойства в глобальной системе моделирования земель, J. Adv. Модель. Earth Sy., 6, 957–979, 2014.
Derksen, C., Xu, X., Scott Dunbar, R., Colliander, A., Kim, Y., Кимбалл, Дж.
С., Блэк, Т. А., Ойскирхен, Э., Ланглуа, А., Лоранти, М. М., Марш, П.,
Раутиайнен, К., Рой, А., Ройер, А., и Стивенс, Дж .: Получение пейзажа
состояние замораживания / оттаивания с радаров с активным пассивным режимом влажности почвы (SMAP) и
радиометрические измерения, Remote Sens. Environ., 194, 48–62,
https://doi.org/10.1016/j.rse.2017.03.007, 2017.
Environ., 194, 48–62,
https://doi.org/10.1016/j.rse.2017.03.007, 2017.
Добсон, М., Улаби, Ф., Халликайнен, М., и Эль-Райес, М .: Микроволновая печь. Диэлектрическое поведение влажной почвы – Часть II: Модели смешивания диэлектриков, IEEE T. Geosci.Пульт, ГЭ-23, 35–46, https://doi.org/10.1109/TGRS.1985.289498, 1985.
Энгстром, Р., Хоуп, А., Квон, Х., Стоу, Д., и Замолодчиков, Д .: Пространственный распределение приповерхностной влажности почвы и ее связь с микрорельеф арктической прибрежной равнины Аляски // Hydrol. Res., 36, 219–234, https://doi.org/10.2166/nh.2005.0016, 2005.
Ойскирхен, Э. С., Брет-Харт, М. С., Шейвер, Г. Р., Эдгар, К. В. и Романовский, В.Е .: Долгосрочный выброс углекислого газа из арктической тундры. Экосистемы на Аляске, Экосистемы, 20, 960–974, https: // doi.org / 10.1007 / s10021-016-0085-9, 2017.
Фаруки, О. Т .: Тепловые свойства почв, Отчет № 81, CRREL.
Монография, Инженерный корпус армии США, Исследовательская и инженерная лаборатория холодных регионов, Ганновер, Нью-Гэмпшир, США, 1981.
Фрей, А., Тедеско, М., Ли, С., Фостер, Дж., Холл, Д.К., Келли Р. и Робинсон, Д. А .: Обзор глобальной снежной продукции, получаемой со спутников. Adv. Космические исследования, 50, 1007–1029, https://doi.org/10.1016/j.asr.2011.12.021, 2012.
Гафуров, А.и Bárdossy, A .: Методика удаления облаков из снежного покрова MODIS. продукт, Hydrol. Earth Syst. Sci., 13, 1361–1373, https://doi.org/10.5194/hess-13-1361-2009, 2009.
Геларо, Р., Маккарти, В., Суарес, М. Дж., Тодлинг, Р., Молод, А., Такач, Л., Рэндлс, К. А., Дарменов А., Босилович М. Г., Райхл Р., Варган К., Кой Л., Куллатер Р., Дрейпер К., Акелла, С., Бучард, В., Конати, А., да Силва, А. М., Гу, В., Ким, Г.-К., Костер, Р., Луччези, Р., Меркова, Д., Нильсен, Дж. Э., Партика, Г., Поусон, С., Путман, В., Ринекер, М., Шуберт, С. Д., Сенкевич, М., и Чжао, Б.: Ретроспективный анализ современной эпохи для исследований и Applications, Version 2 (MERRA-2), J. Climate, 30, 5419–5454, 2017.
Gisnås, K., Westermann, S. , Schuler, T. V., Melvold, K., Etzelmüller, B .:
Мелкомасштабные вариации снега в региональной модели вечной мерзлоты, Криосфера, 10, 1201–1215, https://doi.org/10.5194/tc-10-1201-2016, 2016.
, Schuler, T. V., Melvold, K., Etzelmüller, B .:
Мелкомасштабные вариации снега в региональной модели вечной мерзлоты, Криосфера, 10, 1201–1215, https://doi.org/10.5194/tc-10-1201-2016, 2016.
Гросс, Г., Харден, Дж., Турецкий, М., Макгуайр, А.Д., Камилл, П., Тарноцай, К., Фролкинг, С., Шур, Э.А.Г., Йоргенсон, Т., Марченко, С., Романовский, В., Викленд, К. П., Френч, Н., Уолдроп, М., Буржо-Чавес, Л. и Стригль Р.Г .: Уязвимость почвенного органического углерода в высоких широтах. в Северной Америке к возмущению, J. Geophys. Res.-Biogeo., 116, G00K06, 2011.
Grünewald, T., Bühler, Y., and Lehning, M .: Зависимость от высоты высота снежного покрова в горах, Криосфера, 8, 2381–2394, https://doi.org/10.5194/tc-8-2381-2014, 2014.
Холл, Д.К. и Риггс, Г. А .: Снежный покров MODIS / Terra 8-Day L3 Global 500 м
Сетка, версия 6, Национальные данные по снегу и льду НАСА
Центр Центр распределенного активного архива, Боулдер, Колорадо, США, https://doi.org/10.5067/MODIS/MOD10A2. 006, 2016.
006, 2016.
Хенн, Б., Ньюман, AJ, Ливне, Б., Дейли, К. , и Лундквист, JD: An оценка различий в массивах данных об осадках с координатной привязкой в комплексе местность, J. Hydrol., 556, 1205–1219, https://doi.org/10.1016/j.jhydrol.2017.03.008, 2018.
Хоссейн, М.Ф., Чен В. и Чжан Ю.: Насыпная плотность минеральных и органических веществ. почвы в арктических и субарктических районах Канады, Обработка информации в Agriculture, 2, 183–190, 2015.
Hugelius, G., Strauss, J., Zubrzycki, S., Harden, J. W., Schuur, E. A. G., Ping, C.-L., Ширрмейстер, Л., Гросс, Г., Майклсон, Дж. Дж., Ковен, К. Д., О’Доннелл, Дж. А., Элберлинг, Б., Мишра, У., Камилл, П., Ю, З., Палмтаг, Дж., И Кухри, П .: Оценка запасы углерода циркумполярной вечной мерзлоты с определенными диапазонами неопределенности и выявленные пробелы в данных, Biogeosciences, 11, 6573–6593, https: // doi.org / 10.5194 / bg-11-6573-2014, 2014.
IPCC: Climate Change 2013: The Physical Science Basis, Кембридж
University Press, Кембридж, Соединенное Королевство и Нью-Йорк, Нью-Йорк, США, 1535 стр. , Https://doi.org/10.1017/CBO9781107415324, 2013.
, Https://doi.org/10.1017/CBO9781107415324, 2013.
Джафаров, EE, Coon, ET, Harp, DR, Wilson, CJ , Художник, SL, Этчли, А. Л., Романовский, В. Э .: Моделирование роли предпочтительного снега. накопление через развитие талика и сток грунтовых вод на склонах в переходный ландшафт вечной мерзлоты, Environ.Res. Lett., 13, 105006, https://doi.org/10.1088/1748-9326/aadd30, 2018.
Джин, С. М., Янг, Л. М., Дэниэлсон, П., Гомер, К., Фрай, Дж., И Сиань, Г.: A комплексный метод обнаружения изменений для обновления национального земельного покрова База данных примерно на 2011 год, Remote Sens. Environ., 132, 159–175, 2013.
Келли, Р. Э., Чанг, А. Т., Цанг, Л., и Фостер, Дж. Л .: прототип AMSR-E глобальная площадь снежного покрова и алгоритм высоты снежного покрова, IEEE T. Geosci. Remote, 41, 230–242, https://doi.org/10.1109/TGRS.2003. 809118, 2003.
Ким, Ю., Кимбалл, Дж. С., Робинсон, Д. А., и Дерксен, К.: Новый спутник
записи климатических данных указывают на сильную связь между недавним сезоном заморозков
изменения и снежный покров над высокими северными широтами, Environ. Res. Lett.,
10, 084004, https://doi.org/10.1088/1748-9326/10/8/084004, 2015.
Res. Lett.,
10, 084004, https://doi.org/10.1088/1748-9326/10/8/084004, 2015.
Кинг, Дж., Дерксен, К., Туз, П., Ланглуа, А., Ларсен, К., Лемметинен, Дж., Марш П., Монпети Б., Рой А., Раттер Н. и Штурм М.: Влияние микроструктуры снега по двухчастотным радиолокационным измерениям в тундре окружающая среда, Remote Sens.Окружающая среда., 215, 242–254, https://doi.org/10.1016/j.rse.2018.05.028, 2018.
Кирхнер, П. Б., Бейлс, Р. К., Молотч, Н. П., Фланаган, Дж., и Гуо, Q .: Измерение LiDAR сезонного снегонакопления по перепаду высот в южной Сьерра-Невада, Калифорния, Hydrol. Earth Syst. Sci., 18, 4261–4275, https://doi.org/10.5194/hess-18-4261-2014, 2014.
Киттлер, Ф., Хейманн, М., Колле, О., Зимов, Н. , Зимов, С., и Гёкеде,
М .: Долгосрочный дренаж снижает поглощение CO 2 и выбросы CH 4 в
Экосистема вечной мерзлоты Сибири: влияние дренажа на углеродный цикл Арктики,
Global Biogeochem.Cy., 31, 1704–1717, https://doi.org/10.1002/2017GB005774,
2017.
Ковен, К. Д., Райли, В. Дж., И Стерн, А.: Анализ термического воздействия вечной мерзлоты. динамика и реакция на изменение климата в моделях системы Земли CMIP5, J. Climate, 26, 1877–1900, 2013.
Квок, Р. и Маркус, Т .: Возможные оценки снега в Арктике в масштабе бассейна. глубина с надводным бортом морского льда от CryoSat-2 и ICESat-2: исследовательский анализ, Adv. Космические исследования, 62, 1243–1250, https://doi.org/10.1016/j.asr.2017.09.007, 2018.
Langlois, A., Johnson, C.-A., Montpetit, B., Royer, A., Blukacz-Richards, E. A., Neave, E., Dolant, C., Roy, A., Arhonditsis, G., Kim, D.-K., Kaluskar, С., Брукер, Л .: Обнаружение явлений дождя на снегу (ROS) и слоя льда. формирование с использованием пассивной микроволновой радиометрии: контекст для карибу Пири среда обитания в канадской Арктике, Remote Sens. Environ., 189, 84–95, https://doi.org/10.1016/j.rse.2016.11.006, 2017.
Лоуренс Д. М. и Слейтер А. Г. Включение органических почв в глобальную
климатическая модель, Клим. Dynam., 30, 145–160, 2008.
Dynam., 30, 145–160, 2008.
Лоуренс, Д. М. и Слейтер, А. Г.: Вклад тенденций состояния снега к климату почвы будущего, Клим. Dynam., 34, 969–981, 2010.
Лоуренс, Д. М., Ковен, К. Д., Свенсон, С. К., Райли, В. Дж., И Слейтер, А. G .: Таяние вечной мерзлоты и связанные с этим изменения влажности почвы регулируют прогнозируемые выбросы CO 2 и CH 4 в высоких широтах, Environ. Res. Lett., 10, 094011, https://doi.org/10.1088/1748-9326/10/9/094011, 2015.
Letts, M. G., Roulet, N.Т., Комер, Н. Т., Скарупа, М. Р., Версеги, Д. Л .: Параметризация гидравлических свойств торфяников на территории Канады. схема поверхности, Атмосфера-Океан, 38, 141–160, https://doi.org/10.1080/07055900.2000.9649643, 2000.
Лильедаль, А.К., Бойке, Дж., Даанен, Р.П., Федоров, А.Н., Фрост, Г.В.,
Гросс, Г., Хинзман, Л. Д., Иийма, Ю., Йоргенсон, Дж. К., Матвеева, Н.,
Necsoiu, M., Raynolds, M. K., Romanovsky, V. E., Schulla, J., Tape, K. D.,
Уокер, Д. А., Уилсон, К. Дж., Ябуки, Х., и Зона, Д . : Панарктический ледяной клин.
деградация потепления вечной мерзлоты и ее влияние на гидрологию тундры,
Nat. Geosci., 9, 312–319, 2016.
: Панарктический ледяной клин.
деградация потепления вечной мерзлоты и ее влияние на гидрологию тундры,
Nat. Geosci., 9, 312–319, 2016.
Листон Г. Э. и Штурм М.: Режимы зимних осадков в Арктической Аляске. Определено на основе модели метель и наблюдений за глубиной снега, J. Гидрометеорология, 3, 646–659, https://doi.org/10.1175/1525-7541(2002)003<0646:WPPIAA>2.0.CO;2, 2002.
Лю, С., Вэй, Ю., Пост, ВМ, Повар, РБ, Шефер К. и Торнтон М. М.: Единая карта почв Северной Америки и ее влияние на органический углерод почвы фондовые ресурсы в Северной Америке, Biogeosciences, 10, 2915–2930, https: // doi.org / 10.5194 / bg-10-2915-2013, 2013.
Миронов В. Л., Де Ру Р. Д., Савин И. В. Температурно-зависимый Микроволновая диэлектрическая модель арктической почвы, IEEE T. Geosci. Пульт, 48, 2544–2556, г. https://doi.org/10.1109/TGRS.2010.2040034, 2010.
Мишра, У., Ястроу, Дж. Д., Матамала, Р., Хугелиус, Г., Ковен, К. Д.,
Харден, Дж. У., Пинг, К. Л., Майклсон, Дж. Дж., Фан, З. , Миллер, Р. М.,
Макгуайр, А. Д., Тарнокай, К., Кухри, П., Райли, В. Дж., Шефер, К., Шур,
Э.А.Г., Йоргенсон М.Т., Хинзман Л.Д .: Эмпирические оценки
снизить неопределенности моделирования почвенного органического углерода в районах вечной мерзлоты:
обзор последних достижений и остающихся проблем, Environ. Res. Lett., 8, 035020, г.
2013.
, Миллер, Р. М.,
Макгуайр, А. Д., Тарнокай, К., Кухри, П., Райли, В. Дж., Шефер, К., Шур,
Э.А.Г., Йоргенсон М.Т., Хинзман Л.Д .: Эмпирические оценки
снизить неопределенности моделирования почвенного органического углерода в районах вечной мерзлоты:
обзор последних достижений и остающихся проблем, Environ. Res. Lett., 8, 035020, г.
2013.
Мишра, У., Древняк, Б., Ястроу, Дж. Д., Матамала, Р. М., и Витарана, У. У. А .: Пространственное представление органического углерода и толщины активного слоя высокой широтные почвы в моделях земной системы CMIP5, Geoderma, 300, 55–63, 2016.
Moghaddam, M., Saatchi, S., и Куэнка, Р. Х .: Оценка субскального грунта. влажность с радаром, J. Geophys. Рес.-Атмос., 105, 14899–14911, https://doi.org/10.1029/2000JD
8, 2000. Могхаддам, М., Энтехаби, Д., Гойхман, Ю., Ли, К., Лю, М., Махаджан, А.,
Найяр, А., Шуман, Д., и Тенекетзис, Д.: Умный беспроводной почвенный увлажнитель
Sensor Web с использованием оптимального управления на основе физики: концепция и начало
Демонстрации, IEEE J. Sel. Верхний. Appl., 3, 522–535, г.
https://doi.org/10.1109/JSTARS.2010.2052918, 2010.
Sel. Верхний. Appl., 3, 522–535, г.
https://doi.org/10.1109/JSTARS.2010.2052918, 2010.
Морейра, А., Кригер, Г., Хайнсек, И., Папатанассиу, К., Юнис, М., Лопес-Деккер, П., Хубер, С., Виллано, М., Пардини, М., Эйнедер, М., Де Зан, Ф. и Паризи А .: Tandem-L: инновационная миссия с бистатическим поиском и поиском Глобальное наблюдение динамических процессов на поверхности Земли, IEEE Geosci. Remote Sens. Mag., 3, 8–23, https://doi.org/10.1109/MGRS.2015.2437353, 2015.
Морс П. Д., Берн К. Р. и Кокель С. В. Влияние снега на приповерхностные температуры на возвышенностях и в аллювиальных средах внешняя Дельта Маккензи, Северо-Западные территории, под редакцией: Аллард, М., Жестяная банка. J. Earth Sci., 49, 895–913, https://doi.org/10.1139/e2012-012, 2012.
Наэйми, В., Паулик, К., Барч, А., Вагнер, В., Кидд, Р., Парк, С.-Э. ,
Элгер К. и Бойке Дж .: Флаг состояния поверхности ASCAT (SSF): извлечение
Информация об условиях замораживания / оттаивания поверхности из данных обратного рассеяния с использованием
Эмпирический алгоритм порогового анализа, IEEE T. Geosci.
Remote, 50, 2566–2582, https://doi.org/10.1109/TGRS.2011.2177667, 2012.
Geosci.
Remote, 50, 2566–2582, https://doi.org/10.1109/TGRS.2011.2177667, 2012.
Никольский Д. Дж., Романовский В. Е., Алексеев В. А., Лоуренс Д.М .: Усовершенствованное моделирование динамики вечной мерзлоты в схеме наземной поверхности GCM, Geophys. Res. Lett., 34, L08501, https://doi.org/10.1029/2007GL029525, 2007.
Oechel, W. C., Vourlitis, G., and Hastings, S.J .: Холодное время года CO 2 выбросы из арктических почв // Global Biogeochem. Cy., 11, 163–172, 1997.
Outcalt, S. I., Nelson, F. E., and Hinkel, K. M .: Эффект нулевой завесы: Тепломассообмен через изотермическую область в промерзшем грунте, Вода Ресурс. Res., 26, 1509–1516, https: // doi.org / 10.1029 / WR026i007p01509, 1990.
Пейнтер, Т. Х., Берисфорд, Д. Ф., Бордман, Дж. У., Борман, К. Дж., Димс, Дж.
С., Герке, Ф., Хедрик, А., Джойс, М., Лейдлоу, Р., Маркс, Д., Маттманн,
К., МакГерк, Б., Рамирес, П., Ричардсон, М., Скилс, С. М., Зайдель, Ф. К.,
и Винстрал, А .: Воздушная снежная обсерватория: сочетание сканирующих лидаров,
спектрометр изображений и физическое моделирование для картографирования снежной воды
эквивалент и альбедо снега, Remote Sens. Environ., 184, 139–152,
https: // doi.org / 10.1016 / j.rse.2016.06.018, 2016.
Environ., 184, 139–152,
https: // doi.org / 10.1016 / j.rse.2016.06.018, 2016.
Paquin, J.-P. и Сушама, Л .: Приповерхностная мерзлота Арктики и чувствительность климата к составам моделей почвы и снега в климатических моделях, Клим. Dynam., 44, 203–228, https://doi.org/10.1007/s00382-014-2185-6, 2015.
Parajka, J. и Blöschl, G .: Пространственно-временная комбинация изображений MODIS – потенциал для картографирования снежного покрова: ПРОСТРАНСТВЕННО-ВРЕМЕННАЯ КОМБИНАЦИЯ МОДИСОВ ИЗОБРАЖЕНИЯ, Водные ресурсы. Res., 44, W03406, https://doi.org/10.1029/2007WR006204, 2008.
Parajka, J., Pepe, M., Rampini, A., Rossi, S., and Blöschl, G .: A региональный метод снежного покрова для оценки снежного покрова по MODIS во время облачности обложка, J. Hydrol., 381, 203–212, https://doi.org/10.1016/j.jhydrol.2009.11.042, 2010.
Паразоо, Н. К., Ковен, К. Д., Лоуренс, Д. М., Романовский, В., и Миллер, К. Э .:
Обнаружение углеродной обратной связи вечной мерзлоты: формирование талика и усиление холодного сезона
дыхание как предвестники переходов сток-источник, Криосфера, 12, 123–144, https: // doi.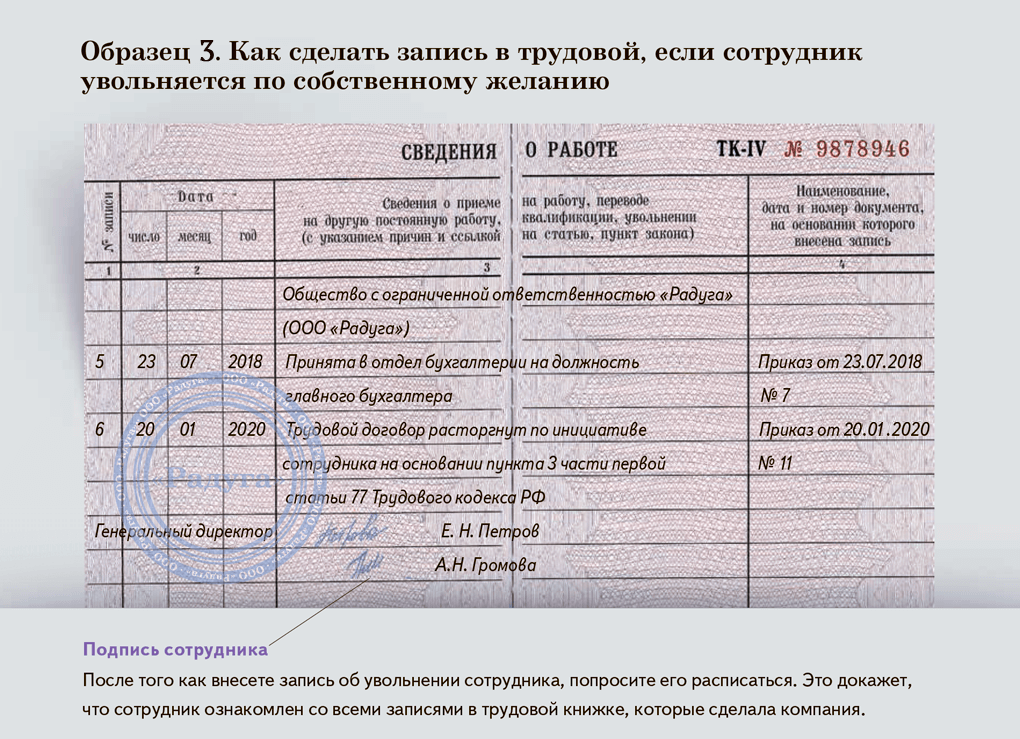 org / 10.5194 / tc-12-123-2018, 2018.
org / 10.5194 / tc-12-123-2018, 2018.
Park, C.-H., Behrendt, A., LeDrew, E., and Wulfmeyer, V: New Approach for Расчет эффективной диэлектрической проницаемости влажной почвы для Микроволны, Remote Sens., 9, 732, https://doi.org/10.3390/rs32, 2017.
Пастик, Н. Дж., Йоргенсон, М. Т., Уайли, Б. К., Нильд, С. Дж., Джонсон, К. Д., Финли А. О .: Распространение приповерхностной вечной мерзлоты на Аляске: Оценки настоящих и будущих условий, Remote Sens. Environ., 168, 301–315, 2015.
Цянь, Б., Грегорич, Э. Г., Гамеда, С., Хопкинс, Д. В., и Ван, X. L .: Наблюдаемые тенденции температуры почвы, связанные с изменением климата в Канаде, J. Geophys. Res., 116, D02106, https://doi.org/10.1029/2010JD015012, 2011.
Роулинз, М. А., Никольский, Д. Дж., Макдональд, К. К., Романовский, В. Э .: Моделирование динамики замерзания / оттаивания почвы с улучшенной панарктической водой модель баланса, J. Adv. Модель. Земля Sy., 5, 659–675, 2013.
Раутиайнен, К., Парккинен, Т. , Лемметьинен, Дж., Шванк, М., Висманн, А.,
Иконен, Дж., Дерксен, К., Давыдов, С., Давыдова, А., Бойке, Дж., Лангер, М.,
Друш М. и Пуллиайнен Дж .: Прототип алгоритма SMOS для обнаружения осени.
замораживание почвы, Remote Sens. Environ., 180, 346–360,
https://doi.org/10.1016/j.rse.2016.01.012, 2016.
, Лемметьинен, Дж., Шванк, М., Висманн, А.,
Иконен, Дж., Дерксен, К., Давыдов, С., Давыдова, А., Бойке, Дж., Лангер, М.,
Друш М. и Пуллиайнен Дж .: Прототип алгоритма SMOS для обнаружения осени.
замораживание почвы, Remote Sens. Environ., 180, 346–360,
https://doi.org/10.1016/j.rse.2016.01.012, 2016.
Райхле, Р. Х., Де Ланнуа, Дж. Дж. М., Лю, К., Ардиццоне, Дж. В., Коллиандер, А., Конати, А.,
Кроу, У., Джексон, Т. Дж., Джонс, Л. А., Кимбалл, Дж. С., Костер, Р. Д., Маханама, С. П.,
Смит, Э. Б., Берг, А., Бирчер, С., Босх, Д., Колдуэлл, Т. Г., Кош, М.,
Гонсалес-Самора, А., Холифилд Коллинз, К. Д., Йенсен, К. Х., Ливингстон, С.,
Лопес-Баеза, Э., Мартинес-Фернандес, Дж., МакНэрн, Х., Могхаддам, М., Пачеко, А.,
Пелларин, Т., Прюгер, Дж., Роулендсон, Т., Сейфрид, М., Старкс, П., Су, З.,
Тибо, М., Ван дер Вельде, Р., Уокер, Дж., Ву, X. и Цзэн, Ю.: Оценка поверхности SMAP уровня 4 и корневой зоны
Продукт влажности почвы с использованием измерений на месте, J.
Гидрометеорология, 18, 2621–2645, 2017.![]()
Романовский, В.Э. и Остеркамп Т. Э .: Влияние незамерзшей воды на тепло и массообменные процессы в активном слое и вечной мерзлоте, вечной мерзлоте Periglac., 11, 219–239, 2000.
Розен, П., Хенсли, С., Шаффер, С., Эдельштейн, В., Ким, Ю., Кумар, Р., Мисра Т., Бхан Р. и Саги Р.: Миссия НАСА-ИСРО SAR (NISAR) эскизный проект двухдиапазонного радиолокационного прибора, в: 2017 IEEE International Симпозиум по геонаукам и дистанционному зондированию (IGARSS), 3832–3835, IEEE, Fort Уорт, Техас, 2017.
Садеги, М., Tabatabaeenejad, A., Tuller, M., Moghaddam, M., and Jones, S .: Улучшение определения влажности почвы в корневой зоне радара AirMOSS P-Band NASA Алгоритм на основе уравнения Ричардса, Remote Sens., 9, 17, https://doi.org/10.3390/rs
17, 2016.
Шефер, К. и Джафаров, Э .: Параметризация дыхания в замороженных почвы на основе наличия субстрата, Biogeosciences, 13, 1991–2001, https://doi.org/10.5194/bg-13-1991-2016, 2016.
Шефер, Г. Л. и Паецольд, Р. F .: SNOTEL (SNOwpack TELemetry) и SCAN (Почва
Сеть климатического анализа), представленная на Автоматизированной метеорологической станции (AWS)
семинар, 6–10 марта, Линкольн, NE, 2000.
F .: SNOTEL (SNOwpack TELemetry) и SCAN (Почва
Сеть климатического анализа), представленная на Автоматизированной метеорологической станции (AWS)
семинар, 6–10 марта, Линкольн, NE, 2000.
Шур, Э.А.Г., Макгуайр, А.Д., Шадель, К., Гросс, Г., Харден, Дж. У., Хейс, Д. Дж., Хугелиус, Г., Ковен, К. Д., Кухри, П., Лоуренс, Д. М., Натали, С. М., Олефельд, Д., Романовский, В. Э., Шефер, К., Турецкий, М. Р., Трит, К. К., Вонк, Дж. Э .: Изменение климата и углерод вечной мерзлоты. обратная связь, Nature, 520, 171–179, 2015.
Слейтер, А.Г. и Лоуренс, Д.М .: Диагностика нынешней и будущей вечной мерзлоты from Climate Models, J. Climate, 26, 5608–5623, 2013.
Smith, S. L., Riseborough, D. W., Bonnaventure, P. P., and Duchesne, C.: An экорегиональная оценка температуры воздуха и поверхности земли в морозные сезоны в коридоре долины Маккензи, СЗТ, Канада, Холодный Рег. Sci. Technol., 125, 152–161, https://doi.org/10.1016/j.coldregions.2016.02.007, 2016.
Штурм М., Тарас Б. , Листон Г.Э., Дерксен, К., Йонас, Т., и Ли, Дж .:
Оценка водного эквивалента снега с использованием данных о глубине снежного покрова и климатических классов,
Ж. Гидрометеорология, 11, 1380–1394, 2010.
, Листон Г.Э., Дерксен, К., Йонас, Т., и Ли, Дж .:
Оценка водного эквивалента снега с использованием данных о глубине снежного покрова и климатических классов,
Ж. Гидрометеорология, 11, 1380–1394, 2010.
Tabatabaeenejad, A., Burgin, M., Duan, X. Y., and Moghaddam, M .: P-Band Радиолокационное извлечение профиля подземной влажности почвы как второй порядок Полином: первые результаты AirMOSS, IEEE T. Geosci. Remote, 53, 645–658, 2015.
Takala, M., Luojus, K., Pulliainen, J., Derksen, C., Lemmetyinen, J., Кярня, Я.-П., Коскинен, Я., и Бойков, Б .: Оценка северных водный эквивалент снега в полушарии для исследования климата путем ассимиляции данных космических радиометров и наземных измерений, Remote Sens. Environ., 115, 3517–3529, https://doi.org/10.1016/j.rse.2011.08.014, 2011.
Thornton, PE, Бег, SW, и Уайт, Массачусетс: создание поверхностей
ежедневные метеорологические переменные на больших участках сложной местности,
J. Hydrol., 190, 214–251,
https://doi.org/10. 1016/S0022-1694(96)03128-9, 1997.
1016/S0022-1694(96)03128-9, 1997.
Throop, J., Левкович А.Г., Смит С.Л .: Климат и почва. температурные отношения на площадках в непрерывном и прерывистом зоны вечной мерзлоты, север Канады, Кан. J. Earth Sci., 49, 865–876, https://doi.org/10.1139/e11-075, 2012.
Геологическая служба США: цифровые модели рельефа Аляски за 2 секунды дуги (DEM) – Сборник загружаемых данных национальной карты USGS 3DEP, США Геологическая служба, 2017.
Walvoord, M.A. и Kurylyk, B.L .: Гидрологические воздействия оттаивания Permafrost-A Review, зона Вадос J., 15, vzj2016.01.0010, 2016.
Ван, З. С. Х. и Халли, Г.: MOD11A2 MODIS / Terra Land Surface Температура / коэффициент излучения 8-дневный L3 Global 1 км SIN Grid V006, NASA EOSDIS Land Процессы DAAC, https://doi.org/10.5067/MODIS/MOD11A2.006, 2015.
Вестерманн, С., Петер, М., Лангер, М., Швамборн, Г., Ширрмайстер, Л.,
Этцельмюллер Б. и Бойке Дж.: Переходное моделирование тепловых процессов в грунте.
условия по спутниковым данным в дельте реки Лена, Сибирь, Криосфера, 11, 1441–1463, https: // doi. org / 10.5194 / tc-11-1441-2017, 2017.
org / 10.5194 / tc-11-1441-2017, 2017.
Ву, М.К .: Гидрология вечной мерзлоты, Шпрингер-Верлаг, Гейдельберг, Германия, 575 С., 2012.
Е, Х., Янг, Д., и Робинсон, Д .: Зимний дождь на снегу и его ассоциации с температурой воздуха на севере Евразии, Hydrol. Процесс., 22, 2728–2736, https://doi.org/10.1002/hyp.7094, 2008.
Йи, Ю., Кимбалл, Дж. С., Роулинз, М. А., Могхаддам, М., и Ойскирхен, Э. С .: Роль снежного покрова, влияющего на промерзание-оттаивание почв бореально-арктического региона. динамика углерода, Биогеонаука, 12, 5811–5829, https: // doi.org / 10.5194 / bg-12-5811-2015, 2015.
Йи, Ю., Кимбалл, Дж. С., Чен, Р. Х., Могхаддам, М., Райхл, Р. Х., Мишра, У., Зона, Д., и Oechel, W.C .: Характеристика динамики активного слоя вечной мерзлоты и чувствительность к пространственной неоднородности ландшафта на Аляске, Криосфера, 12, 145–161, https://doi.org/10.5194/tc-12-145-2018, 2018.
Йошикава, К. и Хинзман, Л.Д .: Сжимающийся термокарст пруды и
динамика подземных вод в прерывистой вечной мерзлоте возле муниципалитета, Аляска,
Вечная мерзлота Периглак. , 14, 151–160, https://doi.org/10.1002/ppp.451,
2003.
, 14, 151–160, https://doi.org/10.1002/ppp.451,
2003.
Юэ, С. Х., Динардо, С. Дж., Акгирай, А., Уэст, Р., Клайн, Д. У. и старейшина, К .: Поляриметрические радиолокационные станции Ku-диапазона дистанционного зондирования наземного снега. Обложка, IEEE T. Geosci. Удаленная, 47, 3347–3364, https://doi.org/10.1109/TGRS.2009.2022945, 2009.
Чжан Т. Дж .: Влияние сезонного снежного покрова на термический режим: Обзор, Rev. Geophys., 43, RG4002, https://doi.org/10.1029/2004RG000157, 2005.
Zona, D., Джоли, Б., Комман, Р., Линдаас, Дж., Вофси, С. К., Миллер, К. Э., Динардо, С. Дж., Денгел, С., Суини, К., Карион, А., Чанг, Р. Ю.-В., Хендерсон, Дж. М., Мерфи, П. К., Гудрич, Дж. П., Моро, В., Лильедаль, А., Уоттс, Дж. Д., Кимбалл, Дж. С., Липсон, Д. А., Ичел, В. К.: Холодная В бюджете метана арктических тундр преобладают сезонные выбросы P. Natl. Акад. Sci. США, 113, 40–45, https://doi.org/10.1073/pnas.1516017113, 2016.
Frontiers | Загадочное волокно Рейсснера и происхождение хордовых
Введение
Тип Chordata, включая ланцетники (cephalochordates – Amphioxus), морских сквиртов (urochordates) и позвоночных, частично характеризуется наличием сегментированной мускулатуры, фланкирующей фиброзный хорд, определяющий полужесткую переднезаднюю ось, и мышечный постблок. анальный хвост используется для плавания.Другой типичной особенностью хордовых является то, что центральная нервная система имеет форму полой нервной трубки, которая проходит от головы до хвоста и заполнена спинномозговой жидкостью (CSF), которая циркулирует по нервному каналу (NCa) (Рисунок 1; Wicht and Lacalli, 2005). ; Aboitiz, Montiel, 2007; Glover, Fritzsch, 2009; Striedter, Northcutt, 2020). Анатомические и поведенческие инновации, приобретенные этой группой, послужили образцом, по которому позвоночные животные возникли и колонизировали землю. Тем не менее, эволюционное происхождение самих хордовых остается одним из больших нерешенных вопросов эволюционной биологии (Satoh, 2008; Holland et al., 2015). В этом контексте хорда рассматривается многими кардинальными чертами хордовых, будучи вовлеченной в развитие некоторых основных признаков этого типа, особенно хвостовой почки и нервной трубки (Henrique et al., 2015; Sasai et al. , 2021).
анальный хвост используется для плавания.Другой типичной особенностью хордовых является то, что центральная нервная система имеет форму полой нервной трубки, которая проходит от головы до хвоста и заполнена спинномозговой жидкостью (CSF), которая циркулирует по нервному каналу (NCa) (Рисунок 1; Wicht and Lacalli, 2005). ; Aboitiz, Montiel, 2007; Glover, Fritzsch, 2009; Striedter, Northcutt, 2020). Анатомические и поведенческие инновации, приобретенные этой группой, послужили образцом, по которому позвоночные животные возникли и колонизировали землю. Тем не менее, эволюционное происхождение самих хордовых остается одним из больших нерешенных вопросов эволюционной биологии (Satoh, 2008; Holland et al., 2015). В этом контексте хорда рассматривается многими кардинальными чертами хордовых, будучи вовлеченной в развитие некоторых основных признаков этого типа, особенно хвостовой почки и нервной трубки (Henrique et al., 2015; Sasai et al. , 2021).
Рис. 1. Схема основных элементов хордового плана тела, изображающая головастиковидное животное с волокнистой хордой (N) и полой нервной трубкой (NT) на спинной стороне (кавитация показана темно-зеленым цветом) , мускулистый хвост (MU) для плавания и жаберные щели (GS, характерный для других deuterostomes). А, анус; H, сердце; М, уст.
А, анус; H, сердце; М, уст.
В этой статье мы предлагаем дополнительное участие волокна Рейсснера (RF), еще одной очень консервативной, но гораздо менее заметной особенности хордовых, которая, вероятно, сотрудничала с хордой в происхождении этой группы животных. RF – это структура, секретируемая NCa, которая возникает на раннем этапе развития большинства хордовых. Несмотря на филогенетическую сохранность и раннее эмбриональное происхождение, функции RF обсуждаются уже более века.Многие авторы предположили роль этого волокна в химической регуляции CSF и в поддержании NCa, что вместе с его филогенетическим консерватизмом может указывать на участие в ранней эволюции хордовой нервной трубки. Более того, недавние исследования указывают на роль этой структуры как проприоцептивного органа, обеспечивающего необходимый мышечный тонус для поддержки прямой оси тела у эмбриона. На наш взгляд, это и другие свидетельства могут пролить свет на роль RF в эволюции плавательного поведения хордовых, возможно, ключевой поведенческой характеристики этого типа. Здесь мы дадим обзор RF, его сравнительную структуру и различные функции, а также обратимся к сравнительным, клиническим и некоторым недавним экспериментальным результатам, чтобы предположить, что эта структура была связана с определением оси тела и плавательного поведения на раннем этапе. хордовые.
Здесь мы дадим обзор RF, его сравнительную структуру и различные функции, а также обратимся к сравнительным, клиническим и некоторым недавним экспериментальным результатам, чтобы предположить, что эта структура была связана с определением оси тела и плавательного поведения на раннем этапе. хордовые.
Характеристика волокна Рейсснера (RF)
Как уже говорилось, RF представляет собой нерастворимое белковое волокно, которое плавает в спинномозговой жидкости внутри хордовой NCa. Он секретируется специализированными эпендимными клетками в мезэнцефальном-диэнцефальном (M-D) соединении и распространяется к каудальному концу нервной трубки, где он разлагается, и его материал попадает в спинномозговую жидкость менингеальной оболочки и кровоток (Rodríguez et al., 1992, 1998). С тех пор как он был впервые описан Рейсснером (1860) на миноге, он остается загадочным компонентом хордовой нервной трубки, которой приписывают многие функции.
Состав и сравнительная структура
Волокно Рейсснера присутствует в NCa большинства хордовых, в том числе цефалохордовых, хордовых и позвоночных, за исключением постнатальных людей и некоторых других млекопитающих. Он состоит из агломерата эластичных волокон, в основном состоящих из гликопротеина SCO-спондина, который содержит большое количество сиаловой кислоты, что усиливает его адгезионные свойства (Olsson and Wingstrand, 1954; Olsson, 1972; Holmberg and Olsson, 1984; Rodríguez et al. ., 1992; Olsson et al., 1994; Gobron et al., 1999). SCO-спондин – это большая молекула внеклеточного матрикса, содержащая многодоменное расположение, которое включает домены фактора фон Виллебранда, повторы SCO-спондина, домены тромбоспондина (TSR) и повторы рецептора ЛПНП, все компоненты, связанные с адгезией клеток и направлением аксонов (Gobron et al. ., 1999; Meiniel, Meiniel, 2007). Существует сильная молекулярная гомология гена SCO-спондина у разных видов, что позволяет предположить, что этот ген является предком всех хордовых (Gobron et al., 1999, 2000; Meiniel et al., 2008). Тем не менее, SCO-spondin значительно удлиняется у позвоночных за счет добавления повторяющихся TSR доменов, что, вероятно, связано с его все более сложной ролью в обеспечении роста аксонов и дифференцировке нейрональных клеток, среди других функций (Meiniel et al.
Он состоит из агломерата эластичных волокон, в основном состоящих из гликопротеина SCO-спондина, который содержит большое количество сиаловой кислоты, что усиливает его адгезионные свойства (Olsson and Wingstrand, 1954; Olsson, 1972; Holmberg and Olsson, 1984; Rodríguez et al. ., 1992; Olsson et al., 1994; Gobron et al., 1999). SCO-спондин – это большая молекула внеклеточного матрикса, содержащая многодоменное расположение, которое включает домены фактора фон Виллебранда, повторы SCO-спондина, домены тромбоспондина (TSR) и повторы рецептора ЛПНП, все компоненты, связанные с адгезией клеток и направлением аксонов (Gobron et al. ., 1999; Meiniel, Meiniel, 2007). Существует сильная молекулярная гомология гена SCO-спондина у разных видов, что позволяет предположить, что этот ген является предком всех хордовых (Gobron et al., 1999, 2000; Meiniel et al., 2008). Тем не менее, SCO-spondin значительно удлиняется у позвоночных за счет добавления повторяющихся TSR доменов, что, вероятно, связано с его все более сложной ролью в обеспечении роста аксонов и дифференцировке нейрональных клеток, среди других функций (Meiniel et al. , 2008). Фактически, многодоменная структура этого белка, вероятно, была создана перетасовкой доменов разных генных доменов, возможно, до возникновения хордовых (Kawashima et al., 2009).
, 2008). Фактически, многодоменная структура этого белка, вероятно, была создана перетасовкой доменов разных генных доменов, возможно, до возникновения хордовых (Kawashima et al., 2009).
Некоторые находки указывают на существование генов, ортологичных SCO-спондину, у иглокожих и полухордовых (оба представляют сестринскую группу хордовых по мнению некоторых авторов; Рисунок 2) и, возможно, у всех двустворчатых животных, предполагая, что RF-подобный материал (но не полимеризованный RF) секретировался радиальными глиеподобными клетками общего предка Bilateria (Gobron et al., 1999; Meiniel et al., 2008; Машанов и др., 2009; Helm et al., 2017). Arendt et al. (2015) предположили, что RF-материал происходит из секреции мукоцилиарных клеток предковых многоклеточных животных (на примере современных плакозоа), которые перемещают частицы пищи во внеклеточную пищеварительную полость (Arendt et al., 2015).
Рисунок 2. Филогения deuterostomes с указанием точек происхождения персонажей, обсуждаемых в этой статье. Большинство филогенетических анализов относят Deuterostomia к монофилетической группе (как показано здесь), а Chordata и ambulacria – к сестринским группам.Однако некоторые недавние открытия с использованием крупномасштабных генетических баз данных утверждают, что нет никаких доказательств того, что дейтеростомия является монофилетической группой (Kapli et al., 2021). Следовательно, в этом узле отображается знак вопроса. Основные выводы этой статьи согласуются с обоими взглядами.
Большинство филогенетических анализов относят Deuterostomia к монофилетической группе (как показано здесь), а Chordata и ambulacria – к сестринским группам.Однако некоторые недавние открытия с использованием крупномасштабных генетических баз данных утверждают, что нет никаких доказательств того, что дейтеростомия является монофилетической группой (Kapli et al., 2021). Следовательно, в этом узле отображается знак вопроса. Основные выводы этой статьи согласуются с обоими взглядами.
Производственные площадки
На позднем этапе развития позвоночных RF секретируется субкомиссуральным органом (SCO), дорсальным околожелудочковым органом, расположенным ниже задней комиссуры в дорсальной нервной трубке (верхней пластине) в месте соединения MD (дорсальный просомер P1; Nieuwenhuys, 1988; Meiniel и другие., 1996; Грондона и др., 2012; Пуэллес, 2018; Муньос и др., 2019; Диас и Пуэллес, 2020 год; Рисунок 3). SCO имеет сложную гистологическую структуру, содержащую специализированные удлиненные эпендимные клетки с базальным отростком, контактирующим с периваскулярным пространством, которое получает синаптические связи, и апикальным отростком, который достигает центрального канала и секретирует RF, среди других компонентов (Rodríguez et al. , 1992). Секреция SCO включает RF и другие компоненты, такие как транспортер тироидных гормонов транстиретрин, основной фактор роста фибробластов и другие гликопротеины, некоторые из которых растворимы и достигают кровотока через базальный конец эпендимных клеток или CSF через апикальный край клетки.Образование RF на апикальной поверхности SCO включает «упаковку» секретируемых белков дисульфидными связями с образованием нерастворимой нити, которая растет вдоль NCa (Rodríguez et al., 1992; Vio et al., 2008; Kiecker, 2018). Однако SCO – не единственное место производства РФ в эмбриогенезе. У ранних эмбрионов нескольких позвоночных, от рыб до млекопитающих, RF материал сначала секретируется (вентральными) клетками пластинки дна вместе с подобными белками, такими как F-spondin. Позже RF материал секретируется специализированным изгибным органом (FO) вентрального M-D головного изгиба (просомеры P1 и / или P2) и собирается в RF.Только на более поздних стадиях SCO начинает вносить вклад в RF вместе с FO, пока последний не прекращает свою функцию, и только SCO производит RF у взрослого человека (Oksche, 1969; Lichtenfeld et al.
, 1992). Секреция SCO включает RF и другие компоненты, такие как транспортер тироидных гормонов транстиретрин, основной фактор роста фибробластов и другие гликопротеины, некоторые из которых растворимы и достигают кровотока через базальный конец эпендимных клеток или CSF через апикальный край клетки.Образование RF на апикальной поверхности SCO включает «упаковку» секретируемых белков дисульфидными связями с образованием нерастворимой нити, которая растет вдоль NCa (Rodríguez et al., 1992; Vio et al., 2008; Kiecker, 2018). Однако SCO – не единственное место производства РФ в эмбриогенезе. У ранних эмбрионов нескольких позвоночных, от рыб до млекопитающих, RF материал сначала секретируется (вентральными) клетками пластинки дна вместе с подобными белками, такими как F-spondin. Позже RF материал секретируется специализированным изгибным органом (FO) вентрального M-D головного изгиба (просомеры P1 и / или P2) и собирается в RF.Только на более поздних стадиях SCO начинает вносить вклад в RF вместе с FO, пока последний не прекращает свою функцию, и только SCO производит RF у взрослого человека (Oksche, 1969; Lichtenfeld et al. , 1999; Meiniel et al., 2008). ).
, 1999; Meiniel et al., 2008). ).
Рис. 3. Волокно Рейсснера (RF, зеленое) в различных хордовых формах и в процессе развития. (A) В нервной трубке цефалохордовых RF секретируется инфундибулярным органом пластинки дна (FP) (IO) и плавает в заполненном спинномозговой жидкостью нервном или центральном канале (NCa). (B – D) Волокно Рейсснера костистых рыб [ (B) , ранний эмбрион; (C) , средняя ступень; (D) , продвинутый]. Обратите внимание, что у костистых изгибов орган (FO, возможно, гомологичный IO у цефалохордовых) заменяется субкомиссуральным органом (SCO) в кровельной пластине (RP) как секретирующим органом RF. Обратите внимание также на нейроны, контактирующие с CSF (CSF-cn) с аксоном (нижний отросток) и дендритом или ресничкой, которые входят в CSF и контактируют с RF.
Эта последовательность развития, по-видимому, соответствует филогенетической истории продукции RF.У цефалохордовых RF секретируется инфундибулярным органом (IO), расположенным в пластине диэнцефального дна (Olsson and Wingstrand, 1954; Olsson, 1972; Olsson et al. , 1994; Wicht and Lacalli, 2005; Рисунок 3). Основываясь на очень схожих секреторных механизмах и топографическом положении (оба расположены в месте, соответствующем головному изгибу, вероятно, в просомере P1 / P2), эмбриональный FO считается гомологичным цефалохордовым IO (Olsson and Wingstrand, 1954). ; Oksche, 1969; Olsson, 1972; Rodríguez et al., 1992; Olsson et al., 1994; Wicht and Lacalli, 2005). То есть обе структуры могут происходить от структуры, присутствующей у последнего общего предка позвоночных и цефалохордовых.
, 1994; Wicht and Lacalli, 2005; Рисунок 3). Основываясь на очень схожих секреторных механизмах и топографическом положении (оба расположены в месте, соответствующем головному изгибу, вероятно, в просомере P1 / P2), эмбриональный FO считается гомологичным цефалохордовым IO (Olsson and Wingstrand, 1954). ; Oksche, 1969; Olsson, 1972; Rodríguez et al., 1992; Olsson et al., 1994; Wicht and Lacalli, 2005). То есть обе структуры могут происходить от структуры, присутствующей у последнего общего предка позвоночных и цефалохордовых.
Примечательно, что урохордовые личинки также продуцируют RF-подобную структуру, которая у некоторых видов секретируется специализированными фибриногеновыми клетками (FC) в хвостовом нервном канатике, которые сравнивали с цефалохордовыми IO в силу секретирования того же материала (Holmberg и Olsson, 1984). Филогенетический анализ показывает, что урохордовые являются сестринской группой позвоночных, а цефалохордовые представляют собой самую раннюю ветвь этого типа (Bourlat et al. , 2006). Если эта точка зрения верна, то Amphioxus ’IO может быть ближе к предковому органу, секретирующему RF, чем urochordate FC, последний является производной структурой, связанной с сидячим образом жизни урохордовых во взрослом возрасте.
, 2006). Если эта точка зрения верна, то Amphioxus ’IO может быть ближе к предковому органу, секретирующему RF, чем urochordate FC, последний является производной структурой, связанной с сидячим образом жизни урохордовых во взрослом возрасте.
Все три органа (IO, FC и SCO) расположены на стыке M-D у разных видов (как сказано, просомер P1), либо в плите пола (IO, FC), либо в плите крыши (SCO). В последние годы было обнаружено, что несколько генов контролируют развитие SCO и продукцию RF (а также развитие других органов), включая Pax6, SOX3, L1-CAM и гены, связанные с апоптозом (Estivill-Torrús et al., 2001; Рамос и др., 2004; Кармона-Калеро и др., 2009; Ли и др., 2012; Мацумото и др., 2020; Ян и др., 2021). Возможно, что экспрессия генов, участвующих в дифференцировке IO (или FO) в пластине дна соединения M-D, кооптировалась в пластине крыши во время происхождения позвоночных по причинам, пока неизвестным. Если это верно, эти структуры, IO / FO с одной стороны и SCO с другой, могут квалифицироваться как последовательные гомологи друг друга (последовательная гомология – это термин, используемый для повторяющихся структур тела, таких как конечности или волосы; Montiel and Aboitiz, 2018 ). Другими словами, в происхождении позвоночных программа предкового развития, участвующая в вентральной секреции RF, активируется в другом регионе, в дорсальной части соединения M-D. Как уже упоминалось, причины этого перехода остаются загадкой.
Другими словами, в происхождении позвоночных программа предкового развития, участвующая в вентральной секреции RF, активируется в другом регионе, в дорсальной части соединения M-D. Как уже упоминалось, причины этого перехода остаются загадкой.
Гипотезы о функциях РФ
Несмотря на все накопленные знания о РФ, нет ясности в отношении его первичной, фундаментальной функции или адаптивного значения (Rodríguez et al., 1992; Meiniel et al., 2008; Бирс и Граймс, 2021 г.). В связи с биением ресничек эпендимных клеток, RF связана с циркуляцией и ионным гомеостазом CSF, где высокое содержание сиаловой кислоты позволяет ему связывать вещества CSF и транспортировать их по нервному каналу по мере роста волокна. В основном RF может участвовать в транспорте и регуляции моноаминов CSF, особенно адреналина, который регулирует секрецию SCO. Точно так же SCO было предложено участвовать в осморегуляции, выведении натрия, диурезе и приеме воды, хотя эти результаты остаются спорными (Rodríguez et al. , 1992; Муньос и др., 2019).
, 1992; Муньос и др., 2019).
Ранний эмбриогенез: поддержание нервного канала
Гидроцефалия – это клиническое состояние, при котором спинномозговая жидкость накапливается в желудочках головного мозга из-за дефектов нервной трубки, нарушающих кровообращение спинномозговой жидкости. Дисфункция комплекса SCO-RF была связана с гидроцефалией у нескольких видов, включая человека. Иммунологическое повреждение SCO или отсутствие RF приводит к коллапсу церебрального водопровода (NCa в соединении M-D), что блокирует отток спинномозговой жидкости и, как следствие, увеличивает внутрижелудочковое давление в полушариях головного мозга (Overholser et al., 1954; Вио и др., 2000; Pérez-Fígares et al., 2001). У мышей с постнатальной гидроцефалией hyh развивается отсутствие NCa в спинном мозге и стеноз переднего мезэнцефального водопровода, а также нарушение SCO и отсутствие RF (Irigoin et al., 1990). У этого мутанта было обнаружено, что эмбриональная денудация эпендимы начинается с вентральной стороны и прогрессирует дорсально во время развития, до начала постнатальной гидроцефалии. Потеря клеток пластинки дна и отсутствие RF у этого мутанта может вызвать искажение эпендимы, которое разрушается из-за давления, создаваемого паренхимой головного мозга (Jiménez et al., 2001; Wagner et al., 2003).
Потеря клеток пластинки дна и отсутствие RF у этого мутанта может вызвать искажение эпендимы, которое разрушается из-за давления, создаваемого паренхимой головного мозга (Jiménez et al., 2001; Wagner et al., 2003).
Роли в последующем развитии и функциях нейронов
Комплекс SCO-RF не только играет роль в раннем развитии, но также различными способами взаимодействует с нервной системой. Во время развития позвоночных SCO и его секретируемые белки были связаны с направлением аксонов в средней линии нервной трубки, что характерно для верхней и нижней пластин нервной трубки эмбриона (Meiniel et al., 1996; Gobron et al., 2000 ; Grondona et al., 2012).RF также был связан с регенерацией хвостовой нервной трубки у низших позвоночных (Meiniel et al., 1996; Alibardi, 2021), и было предложено, что растворимый RF-материал, секретируемый SCO, способствует нейрогенезу взрослых у млекопитающих (Guerra et al. ., 2015). Кроме того, SCO – это сильно иннервируемый орган, получающий различные входные данные, несущие моноамины, ацетилхолин, нейропептиды и ГАМК. Некоторые данные свидетельствуют о том, что серотонин и другие моноамины могут модулировать секреторную активность SCO, возможно, за счет сигналов, генерируемых самой нервной трубкой (Rodríguez et al., 1992; Richter et al., 2004; Эль-Хиба и др., 2020).
Некоторые данные свидетельствуют о том, что серотонин и другие моноамины могут модулировать секреторную активность SCO, возможно, за счет сигналов, генерируемых самой нервной трубкой (Rodríguez et al., 1992; Richter et al., 2004; Эль-Хиба и др., 2020).
Нейроны, контактирующие с ЦСЖ
Возможно, более актуально для этой статьи, стенки NCa содержат обильные CSF-контактирующие нейроны (CSF-cn) по всей своей длине (Rodríguez et al., 1992; Bearce and Grimes, 2021). CSF-cn проецирует подобные дендритам расширения в NCa и расширяет большую ресничку и несколько стереоцилий, которые контактируют с RF. Хотя некоторые из этих нейронов, по-видимому, являются ГАМКергическими, они также положительны для некоторых других нейротрансмиттеров, включая моноамины, соматостатин (SST) и пептид, родственный уротензину II (URP).Нейроны URP иннервируются аксонами SST +, тогда как нейроны SST + получают серотонинергическую иннервацию (Rodríguez et al., 1992; Bearce and Grimes, 2021). Эти нейроны распространяют аксоны на латеровентральный спинной мозг и др. Регионы, и было предложено обеспечить механизм обратной связи, с помощью которого CSF-cn может контролировать активность SCO (Rodríguez et al., 1992). Таким образом, CSF-cn может воспринимать различные химические и механические стимулы от CSF и RF, включая изгиб RF и оси тела и стимуляцию спинной мускулатуры для коррекции этих отклонений (Böhm et al., 2016; Jalalvand et al., 2016). Последнее предполагает роль CSF-cn и RF в передвижении и положении тела (Gobron et al., 1999, 2000), вопрос, который будет обсуждаться ниже.
Регионы, и было предложено обеспечить механизм обратной связи, с помощью которого CSF-cn может контролировать активность SCO (Rodríguez et al., 1992). Таким образом, CSF-cn может воспринимать различные химические и механические стимулы от CSF и RF, включая изгиб RF и оси тела и стимуляцию спинной мускулатуры для коррекции этих отклонений (Böhm et al., 2016; Jalalvand et al., 2016). Последнее предполагает роль CSF-cn и RF в передвижении и положении тела (Gobron et al., 1999, 2000), вопрос, который будет обсуждаться ниже.
Разработка прямой оси тела
Кроме того, несколько исследований показывают, что RF-изменения участвуют в патогенезе сколиоза. В этом клиническом состоянии позвоночник изгибается в сторону вместо того, чтобы сохранять прямую ось, что влияет на осанку и ухудшается с возрастом. В начале прошлого века Николлс (1913) и Колмер (1921) предположили, что РФ может работать как осевой проприоцептивный орган, обеспечивая соматосенсорный контроль над телом и способствуя поддержанию оси тела на личиночной стадии. Другие исследования, проведенные в прошлом веке, показали, что нарушение SCO или RF вызывает искажение оси тела у личинок земноводных и рыб (см. Rodríguez et al., 1992). Дополнительные данные показали, что нарушение движений ресничек эпендимы нарушает отток спинномозговой жидкости и вызывает искривление позвоночника у рыбок данио, предполагая связь между движением ресничек и функцией RF (Brand et al., 1996; Kramer-Zucker et al., 2005; Bearce and Grimes) , 2021). Более поздние исследования, проведенные на рыбках данио, подтвердили эту возможность, продемонстрировав, что RF имеет решающее значение для поддержания прямой оси тела и морфогенеза позвоночника, поскольку отсутствие RF во время развития приводит к сколиозу у взрослых (Driever, 2018; Ringers and Jurisch-Yaksi , 2020).В первом исследовании Cantaut-Belarif et al. (2018) исключили РФ у мутантов, лишенных SCO-спондина. В первый день после вылупления мутантные личинки имели обычно прямую ось тела (возможно, поддерживаемую формирующейся хордой), но через 30 часов личинки демонстрировали искаженную кривизну туловища.
Другие исследования, проведенные в прошлом веке, показали, что нарушение SCO или RF вызывает искажение оси тела у личинок земноводных и рыб (см. Rodríguez et al., 1992). Дополнительные данные показали, что нарушение движений ресничек эпендимы нарушает отток спинномозговой жидкости и вызывает искривление позвоночника у рыбок данио, предполагая связь между движением ресничек и функцией RF (Brand et al., 1996; Kramer-Zucker et al., 2005; Bearce and Grimes) , 2021). Более поздние исследования, проведенные на рыбках данио, подтвердили эту возможность, продемонстрировав, что RF имеет решающее значение для поддержания прямой оси тела и морфогенеза позвоночника, поскольку отсутствие RF во время развития приводит к сколиозу у взрослых (Driever, 2018; Ringers and Jurisch-Yaksi , 2020).В первом исследовании Cantaut-Belarif et al. (2018) исключили РФ у мутантов, лишенных SCO-спондина. В первый день после вылупления мутантные личинки имели обычно прямую ось тела (возможно, поддерживаемую формирующейся хордой), но через 30 часов личинки демонстрировали искаженную кривизну туловища. Как уже упоминалось, подобный фенотип наблюдался у животных с дефектами движения ресничек в эпендимных клетках, выстилающих нервный канал, что нарушает циркуляцию спинномозговой жидкости (Bearce and Grimes, 2021). Однако цилиогенез и циркуляция CSF нормальны у мутантов SCO-spondin.С другой стороны, у мутантов с дефектами ресничек RF не образуется. Cantaut-Belarif et al. (2018) пришли к выводу, что движение ресничек необходимо для формирования RF и что движение ресничек, но не движение ресничек, необходимо для поддержания прямой оси тела на протяжении личиночного развития. В последующем исследовании гипоморфные мутантные личинки SCO-spondin (экспрессирующие более низкие уровни белка и образующие дефектную RF) вызывали искривленный хвост и сколиоз у взрослых (Troutwine et al., 2020).
Как уже упоминалось, подобный фенотип наблюдался у животных с дефектами движения ресничек в эпендимных клетках, выстилающих нервный канал, что нарушает циркуляцию спинномозговой жидкости (Bearce and Grimes, 2021). Однако цилиогенез и циркуляция CSF нормальны у мутантов SCO-spondin.С другой стороны, у мутантов с дефектами ресничек RF не образуется. Cantaut-Belarif et al. (2018) пришли к выводу, что движение ресничек необходимо для формирования RF и что движение ресничек, но не движение ресничек, необходимо для поддержания прямой оси тела на протяжении личиночного развития. В последующем исследовании гипоморфные мутантные личинки SCO-spondin (экспрессирующие более низкие уровни белка и образующие дефектную RF) вызывали искривленный хвост и сколиоз у взрослых (Troutwine et al., 2020).
Другое исследование продемонстрировало, что RF стимулирует механосенсорные реснички CSF-cn в позвоночном канале.Когда ось прямая, RF плавает в нервном канале, но когда он изгибается, он входит в контакт с механосенсорными нейронами во внутренней стенке нервной трубки, которые активируют осевую мускулатуру, восстанавливая ось жесткости тела (Orts-Del’Immagine et al. , 2020 ). Две следующие статьи пролили дополнительный свет на механосенсорную трансдукцию RF-CSF-cn (Cantaut-Belarif et al., 2020; Lu et al., 2020). Эти исследования показали, что URP требует экспрессии интактного RF и подавляется у мутантов SCO-spondin, что приводит к нарушению передачи сигналов в мышцы туловища.Кроме того, лечение мутантов адреналином увеличивало экспрессию URP и спасало фенотип, предполагая, что этот нейромедиатор опосредует передачу сигналов RF-CSF-cn, возможно, через транзиенты кальция в спинномозговых нейронах. Кроме того, обеспечение URP CSF привело к восстановлению фенотипа личиночных и взрослых мутантов. Наконец, у мутантов SCO-spondin также развиваются повышенные нейровоспалительные реакции, связанные со сколиозом. Подавление воспаления приводит к восстановлению фенотипа дикого типа (Rose et al., 2020). Обсуждая эти результаты, Рингерс и Юриш-Якси (2020) предположили, что RF защищает спинной мозг от воспаления и нарушает нормальное поддержание прямой оси тела, подавляя механосенсорные рефлексы, которые поддерживают жесткую осанку тела.
, 2020 ). Две следующие статьи пролили дополнительный свет на механосенсорную трансдукцию RF-CSF-cn (Cantaut-Belarif et al., 2020; Lu et al., 2020). Эти исследования показали, что URP требует экспрессии интактного RF и подавляется у мутантов SCO-spondin, что приводит к нарушению передачи сигналов в мышцы туловища.Кроме того, лечение мутантов адреналином увеличивало экспрессию URP и спасало фенотип, предполагая, что этот нейромедиатор опосредует передачу сигналов RF-CSF-cn, возможно, через транзиенты кальция в спинномозговых нейронах. Кроме того, обеспечение URP CSF привело к восстановлению фенотипа личиночных и взрослых мутантов. Наконец, у мутантов SCO-spondin также развиваются повышенные нейровоспалительные реакции, связанные со сколиозом. Подавление воспаления приводит к восстановлению фенотипа дикого типа (Rose et al., 2020). Обсуждая эти результаты, Рингерс и Юриш-Якси (2020) предположили, что RF защищает спинной мозг от воспаления и нарушает нормальное поддержание прямой оси тела, подавляя механосенсорные рефлексы, которые поддерживают жесткую осанку тела.
Обсуждение: роль RF в ранней эволюции хордовых
Как мы обсуждали выше, несмотря на то, что проводится много исследований RF, пока нет ясности относительно его первичной роли в развитии нервной системы или в эволюции хордовых.Поскольку RF и SCO необходимы для предотвращения коллапса церебрального водопровода позвоночных, а полимеризованный RF существует только в хордовых, некоторые авторы спрашивают, могли ли эти компоненты (точнее, RF и цефалохордовый IO) способствовать формированию нервной трубки в хордовых началах (Meiniel et al., 2008). Однако полая нервная система не может иметь хордовый характер. Нервный шнур полухордовых (Рис. 2, 4) обнаруживает локализованную кавитацию, которая для многих авторов предвещает хордовую нервную трубку (Kaul and Stach, 2010), хотя есть и другие мнения (Satoh, 2008).В любом случае происхождение нейруляции остается нерешенной проблемой. Раннее предположение предписывало, что нейруляция является результатом инвагинации двух продольных цилиарных полос, присутствующих у гипотетической предковой личинки диплеврулы (Garstang, 1894; Holland, 2011). Гипотеза привлекательна, но пока нет доказательств в ее поддержку. Также возможно, что секреция неполимеризованных белков SCO-спондинов в апикальной нейроэктодерме способствовала ранней инвагинации нервного канатика в ранних дейтеростомах или хордовых (Meiniel et al., 2008), хотя опять же необходимы дополнительные доказательства в этой области.
Гипотеза привлекательна, но пока нет доказательств в ее поддержку. Также возможно, что секреция неполимеризованных белков SCO-спондинов в апикальной нейроэктодерме способствовала ранней инвагинации нервного канатика в ранних дейтеростомах или хордовых (Meiniel et al., 2008), хотя опять же необходимы дополнительные доказательства в этой области.
Рис. 4. Выше нервная трубка хордовой (эмбриональной рыбы) показывает субкомиссуральный орган (SCO) и изгибный орган (FO), места происхождения RF. Эти органы расположены на мезэнцефальной-диэнцефальной границе, которая включена в эмбриональную область, ограниченную zona limitans intrathalamica (ZLI) спереди и границей среднего мозга-заднего мозга (MHB) сзади. Эта область является сильно Otx-положительной на раннем этапе развития и по этой причине считается гомологичной расположению воротниковой области (C) тела полухордовых (ниже), демонстрирует те же морфогенетические границы (гомологи ZLI и MHB) и содержит нервный шнур с полым воротничком (CNC) (Pani et al. , 2012; Lowe et al., 2015). Помимо ЧПУ, полухордовые демонстрируют дорсальные и вентральные нервные тяжи (DNC, VNC) в туловище тела (T) и диффузную нейронную сеть (NNT, заштрихованные области), покрывающую тело (Holland et al., 2015). А, анус; GS, жаберные щели; М, рот; П, хоботок.
, 2012; Lowe et al., 2015). Помимо ЧПУ, полухордовые демонстрируют дорсальные и вентральные нервные тяжи (DNC, VNC) в туловище тела (T) и диффузную нейронную сеть (NNT, заштрихованные области), покрывающую тело (Holland et al., 2015). А, анус; GS, жаберные щели; М, рот; П, хоботок.
Следует отметить, что полухордовая нервная трубка расположена в воротниковой области головы, области, строго положительной для генов Otx и Pax6, чей аналог в хордовых включает MD-соединение, согласно исследованиям картирования экспрессии генов (Рисунок 4; Williams and Голландия, 1996; Gobron et al., 1999; Голландия и Шорт, 2008; Пани и др., 2012; Lowe et al., 2015). Одна возможность состоит в том, что в хордовых источниках сборка RF была связана с развитием расширенного NCa, проходящего не только в передних областях, но также и в каудальных Otx-негативных областях предкового нервного канатика; то есть в туловище и постанальный хвост. Эмбриональное удлинение нервной трубки – фундаментальный процесс, формирующий хордовый план тела (Lacalli, 2000).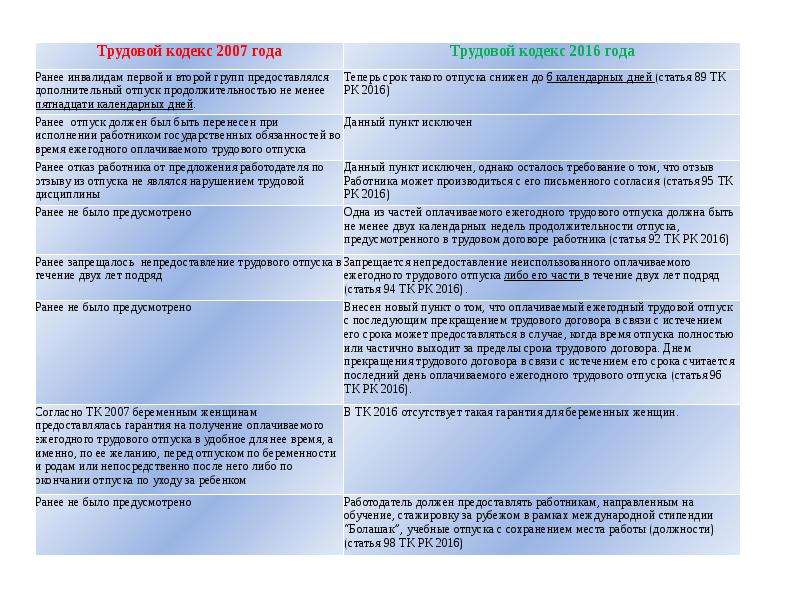 Тем не менее, у низших позвоночных, включая рыб с плавниками и миногу, кавитация нервной трубки, по-видимому, не возникает сразу, но сначала имеется сплошной костномозговой канатик (нервный киль), который кавитирует на разных уровнях по своей длине, превращаясь в непрерывный нервный канал. только позже в развитии (Handrigan, 2003).Напоминая о роли RF в предотвращении гидроцефалии, эта структура могла участвовать в производстве и поддержании непрерывных NCa вдоль продольной оси тела у этих видов. Кроме того, интересно напомнить, что задний спинной мозг, иннервирующий хордовый хвост, производится вторичной нейруляцией, т. Е. Образованием вторичной нервной трубки, происходящей из мезенхимы хвостовой почки, которая позже сливается с главной нервной трубкой (Handrigan, 2003; Beaster-Jones et al., 2008; Энрике и др., 2015). У головохордовых, хордовых и позвоночных NCa хвоста содержит RF (Obermüller-Wilén and Olsson, 1974), а RF также присутствует в регенерирующем хвосте лепидозавров и других позвоночных (Meiniel et al.
Тем не менее, у низших позвоночных, включая рыб с плавниками и миногу, кавитация нервной трубки, по-видимому, не возникает сразу, но сначала имеется сплошной костномозговой канатик (нервный киль), который кавитирует на разных уровнях по своей длине, превращаясь в непрерывный нервный канал. только позже в развитии (Handrigan, 2003).Напоминая о роли RF в предотвращении гидроцефалии, эта структура могла участвовать в производстве и поддержании непрерывных NCa вдоль продольной оси тела у этих видов. Кроме того, интересно напомнить, что задний спинной мозг, иннервирующий хордовый хвост, производится вторичной нейруляцией, т. Е. Образованием вторичной нервной трубки, происходящей из мезенхимы хвостовой почки, которая позже сливается с главной нервной трубкой (Handrigan, 2003; Beaster-Jones et al., 2008; Энрике и др., 2015). У головохордовых, хордовых и позвоночных NCa хвоста содержит RF (Obermüller-Wilén and Olsson, 1974), а RF также присутствует в регенерирующем хвосте лепидозавров и других позвоночных (Meiniel et al. , 1996; Alibardi, 2021). Следовательно, было бы очень интересно исследовать, вносит ли RF вклад в удлинение и поддержание вторичного нервного канала в задней почке ранних хордовых.
, 1996; Alibardi, 2021). Следовательно, было бы очень интересно исследовать, вносит ли RF вклад в удлинение и поддержание вторичного нервного канала в задней почке ранних хордовых.
Помимо своей возможной роли в раннем эмбриогенезе хордовых, RF, возможно, также был критическим для развития поведения хордового плавания, что в конечном итоге оказалось ключом к успеху позвоночных.Ввиду недавних данных, указывающих на роль RF в развитии прямой оси, мы предварительно предполагаем, что у ранних хордовых наличие механически пассивной хорды могло быть недостаточным для поддержания жесткости переднезадней оси в качестве эмбриона. вырос в размерах. Для поддержания прямой формы тела требовалась стимуляция сократительных элементов хорды и спинной мускулатуры, что обеспечивалось проприоцептивной схемой RF / CSF-cn. Другими словами, RF в контакте с CSF-cn (экспрессирующим URP или другими нейротрансмиттерами), возможно, обеспечил датчик положения тела, который поддерживал необходимую плотность хорды и спинной мускулатуры у растущей личинки, максимизируя упругую энергию во время плавательных гребков. Подводя итог, можно сказать, что удлинение NCa в области туловища и хвоста, содержащих внутреннюю RF в контакте с CSF-cn, могло быть ключевым приобретением ранних хордовых, которые позволили не только поддерживать прямую ось, но и, возможно, также способствовали нервной системе. контроль, необходимый для эффективного плавания.
Подводя итог, можно сказать, что удлинение NCa в области туловища и хвоста, содержащих внутреннюю RF в контакте с CSF-cn, могло быть ключевым приобретением ранних хордовых, которые позволили не только поддерживать прямую ось, но и, возможно, также способствовали нервной системе. контроль, необходимый для эффективного плавания.
Примечательно, что недавние исследования пролили свет на другое высококонсервативное событие в раннем развитии хордовых: небольшое вентральное изгибание эмбрионального хвоста во время раннего развития, чтобы соответствовать кривизне эмбриональных мембран.Хотя это считалось результатом внешних сил, воздействующих на растущий эмбрион, было обнаружено, что актомиозин накапливается на вентральной стороне хорды, условие, которое необходимо для правильного изгиба хвоста (Lu et al., 2020). Участвуют ли RF и CSF-cn в этом процессе – также вопрос дальнейших исследований.
Заключительные комментарии
В этой статье рассмотрены некоторые старые и недавние открытия относительно функции RF с эволюционной точки зрения. Мы предполагаем, что сборка RF в виде растущих филаментов в рудиментарных NCa хордовых предков вносит вклад в расширение нервной трубки в область хвоста одновременно с дифференцировкой хорды.Это нововведение также обеспечило проприоцептивный контроль туловища и хвоста, облегчая поддержание прямой оси и развитие сократительного плавательного хвоста. RF был впервые обнаружен около 160 лет назад, и его функция и строгий консерватизм для всех хордовых остались загадкой эволюции. Недавние открытия могут дать критическое понимание его основной функции во время раннего развития хордовых и их сохранения в филюме. Дальнейшие исследования, направленные на раскрытие возможного участия РФ или его неполимеризованных компонентов в процессе нейруляции, а также роли этой структуры в развитии оси тела и плавательного поведения могут пролить некоторый свет на происхождение этой интригующей компонент нервной трубки.Наконец, понимание роли РФ в нервном развитии может способствовать лечению заболеваний спинного мозга, включая гидроцефалию и сколиоз.
Мы предполагаем, что сборка RF в виде растущих филаментов в рудиментарных NCa хордовых предков вносит вклад в расширение нервной трубки в область хвоста одновременно с дифференцировкой хорды.Это нововведение также обеспечило проприоцептивный контроль туловища и хвоста, облегчая поддержание прямой оси и развитие сократительного плавательного хвоста. RF был впервые обнаружен около 160 лет назад, и его функция и строгий консерватизм для всех хордовых остались загадкой эволюции. Недавние открытия могут дать критическое понимание его основной функции во время раннего развития хордовых и их сохранения в филюме. Дальнейшие исследования, направленные на раскрытие возможного участия РФ или его неполимеризованных компонентов в процессе нейруляции, а также роли этой структуры в развитии оси тела и плавательного поведения могут пролить некоторый свет на происхождение этой интригующей компонент нервной трубки.Наконец, понимание роли РФ в нервном развитии может способствовать лечению заболеваний спинного мозга, включая гидроцефалию и сколиоз. Кроме того, учитывая возможное участие RF в удлинении нервного канала и регенерации хвоста у низших позвоночных, дальнейшие исследования могут быть направлены на раскрытие терапевтического участия этой структуры в регенерации спинного мозга после травмы или дегенеративного заболевания.
Кроме того, учитывая возможное участие RF в удлинении нервного канала и регенерации хвоста у низших позвоночных, дальнейшие исследования могут быть направлены на раскрытие терапевтического участия этой структуры в регенерации спинного мозга после травмы или дегенеративного заболевания.
Взносы авторов
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Aboitiz, F., and Montiel, J. (2007). Происхождение и эволюция конечного мозга позвоночных, с особым упором на неокортекс млекопитающих. Adv. Анат. Эмбриол. Ячейка Биол . 193, 1–112. DOI: 10.1007 / 978-3-540-49761-5
CrossRef Полный текст | Google Scholar
Алибарди, Л.(2021 г. ). Регенерация хвоста у лепидозавров как исключение из общего отсутствия регенерации органов у амниот. Exp. Zool. B. Mol. Dev. Evol. 336, 145–164. DOI: 10.1002 / jez.b.22901
). Регенерация хвоста у лепидозавров как исключение из общего отсутствия регенерации органов у амниот. Exp. Zool. B. Mol. Dev. Evol. 336, 145–164. DOI: 10.1002 / jez.b.22901
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Арендт, Д., Бенито-Гутьеррес, Э., Брюне, Т., и Марлоу, Х. (2015). Желудочные мешки и мукоцилиарная подошва: почва для эволюции нервной системы. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 370: 20150286.DOI: 10.1098 / rstb.2015.0286
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бирс, Э. А., Граймс, Д. Т. (2021 г.). О правильной форме: роли подвижных ресничек и спинномозговой жидкости в морфологии тела и позвоночника. Семин. Cell Dev. Биол . 110, 104–112. DOI: 10.1016 / j.semcdb.2020.07.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бистер-Джонс, Л., Кальтенбах, С. Л., Куп, Д., Юань, С., Честейн, Р., и Холланд, Л. З. (2008). Экспрессия генов сегментации сомитов в амфиоксусе: часы без волнового фронта? Dev. Genes Evol. 218, 599–611. DOI: 10.1007 / s00427-008-0257-5
Genes Evol. 218, 599–611. DOI: 10.1007 / s00427-008-0257-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бём У. Л., Прендергаст А., Дженун Л., Нунес Фигейредо С., Гомес Дж., Стокс К. и др. (2016). Нейроны, контактирующие с ЦСЖ, регулируют движение, передавая механические стимулы в спинномозговые цепи. Nat. Commun. 7: 10866.DOI: 10.1038 / ncomms10866
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бурлат С. Дж., Юлиусдоттир Т., Лоу К. Дж., Фриман Р., Аронович Дж., Киршнер М. и др. (2006). Филогения Deuterostome выявляет монофилетические хордовые и новый тип Xenoturbellida. Природа 444, 85–88. DOI: 10.1038 / nature05241
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бранд, М., Гейзенберг, К. П., Варга, Р. М., Пелегри, Ф., Karlstrom, R.O., Beuchle, D., et al. (1996). Мутации, влияющие на развитие средней линии и общей формы тела во время эмбриогенеза рыбок данио. Разработка 123, 129–142.
Google Scholar
Кантаут-Белариф, Ю., Ортс Дель’Иммаджин, А., Пенру, М., Пезерон, Г., Вярт, К., и Барде, П. Л. (2020). Адренергическая активация модулирует сигнал от волокна Рейсснера к нейронам, контактирующим с спинномозговой жидкостью, во время развития. Элиф 9: e59469. DOI: 10.7554 / eLife.59469
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кантаут-Белариф, Ю., Штернберг, Дж. Р., Тувенин, О., Вярт, К., и Барде, П. Л. (2018). Волокно Рейсснера в спинномозговой жидкости контролирует морфогенез оси тела. Curr. Биол. 28, 2479.e4–2486.e4.
Google Scholar
Кармона-Калеро, Э. М., Гонсалес-Марреро, И., Гонсалес-Толедо, Дж. М., Кастаньейра-Руис, А., Де Пас-Кармона, Х., Кастаньейра-Руис, Л. и др.(2009). Белки клетчатки Рейсснера и вариации p73 в спинномозговой жидкости и субкомиссуральном органе гидроцефальных крыс. Анат. Histol. Эмбриол . 38, 282–285. DOI: 10.1111 / j.1439-0264.2009.00939.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эль-Хиба, О. , Драуи, А., и Гамрани, Х. (2020). Нейроактивный нейростероид дегидроэпиандростерон сульфат (DHEAS) модулирует серотонинергическую систему в дорсальном ядре Raphe и высвобождение спинномозговой жидкости волокна Рейсснера у крыс. С. Р. Биол . 343, 101–110. DOI: 10.5802 / crbiol.3
, Драуи, А., и Гамрани, Х. (2020). Нейроактивный нейростероид дегидроэпиандростерон сульфат (DHEAS) модулирует серотонинергическую систему в дорсальном ядре Raphe и высвобождение спинномозговой жидкости волокна Рейсснера у крыс. С. Р. Биол . 343, 101–110. DOI: 10.5802 / crbiol.3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эстивиль-Торрус, Г., Виталис, Т., Фернандес-Ллебрес, П., и Прайс, Д. Дж. (2001). Фактор транскрипции Pax6 необходим для развития диэнцефальной дорсальной срединной секреторной радиальной глии, которая формирует субкомиссуральный орган. мех. Dev . 109, 215–224. DOI: 10.1016 / s0925-4773 (01) 00527-5
CrossRef Полный текст | Google Scholar
Гарстанг, W.(1894). Предварительное замечание о новой теории филогении хордовых. Zool. Анз . 17, 122–125.
Google Scholar
Гловер, Дж. К., и Фриц, Б. (2009). «Мозги примитивных хордовых», в Encyclopedia of Neuroscience , 4-е изд. , Vol. 6, изд. Л. Сквайр (Оксфорд: Academic Press), 439–448.
, Vol. 6, изд. Л. Сквайр (Оксфорд: Academic Press), 439–448.
Google Scholar
Гоброн, С., Крево, И., Мейниэль, Р., Дидье, Р., Дастуг, Б., и Мейниэль, А. (1999). SCO-спондин эволюционно консервативен в центральной нервной системе типа хордовых. Неврология 88, 655–664. DOI: 10.1016 / s0306-4522 (98) 00252-8
CrossRef Полный текст | Google Scholar
Gobron, S., Creveaux, I., Meiniel, R., Didier, R., Herbet, A., Bamdad, M., et al. (2000). Подкомиссуральный орган / комплекс волокон Рейсснера: характеристика SCO-спондина, гликопротеина с сильным действием на рост нейритов. Glia 32, 177–191. DOI: 10.1002 / 1098-1136 (200011) 32: 2 <177 :: aid-glia70 <3.0.co; 2-v
CrossRef Полный текст | Google Scholar
Грондона, Дж.М., Хойо-Бесерра, К., Виссер, Р., Фернандес-Ллебрес, П., и Лопес-Авалос, М. Д. (2012). Подкомиссуральный орган и развитие задней комиссуры. Внутр. Rev. Cell Mol. Биол . 296, 63–137. DOI: 10.1016 / B978-0-12-394307-1. 00002-3
00002-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Герра М. М., Гонсалес К., Каприле Т., Хара М., Вио К., Муньос Р. И. и др. (2015). Понимание того, как субкомиссуральный орган и другие перивентрикулярные секреторные структуры вносят вклад через спинномозговую жидкость в нейрогенез. Перед. Яковлевск . 9: 480. DOI: 10.3389 / fncel.2015.00480
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хелм, К., Карл, А., Бекерс, П., Кауль-Штрелов, С., Ульбрихт, Э., Куртезис, И., и др. (2017). Ранняя эволюция радиальных глиальных клеток Bilateria. Proc. Биол. Sci. 284: 20170743. DOI: 10.1098 / rspb.2017.0743
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энрике, Д., Абранш, Э., Верриер, Л., и Стори, К. Г. (2015). Нейромезодермальные предшественники и формирование спинного мозга. Развитие 142, 2864–2875. DOI: 10.1242 / dev.119768
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холланд, Л. З., и Шорт, С. (2008). Дупликация, кооптация и рекрутирование генов при происхождении мозга позвоночных от хордового мозга беспозвоночных. Brain Behav. Evol . 72, 91–105. DOI: 10.1159 / 000151470
З., и Шорт, С. (2008). Дупликация, кооптация и рекрутирование генов при происхождении мозга позвоночных от хордового мозга беспозвоночных. Brain Behav. Evol . 72, 91–105. DOI: 10.1159 / 000151470
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Голландия, Н.Д., Холланд, Л. З., и Холланд, П. У. (2015). Сценарии создания позвоночных. Природа 520, 450–455.
Google Scholar
Холмберг, К., и Олссон, Р. (1984). Происхождение волокна Рейсснера от аппендикуляра Oikopleura dioica. Виденск. Meddr Dansk Naturh. Форен . 145, 43–52.
Google Scholar
Иригоин, К., Родригес, Э. М., Хайнрихс, М., Фрезе, К., Херцог, С., Окше, А., и др. (1990). Иммуноцитохимическое исследование подкомиссурального органа крыс с индуцированной постнатальной гидроцефалией. Exp. Мозг Res . 82, 384–392. DOI: 10.1007 / BF00231257
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джалалванд, Э., Робертсон, Б. , Валлен, П., и Грилльнер, С. (2016). Ресничные нейроны, выстилающие центральный канал, воспринимают движение жидкости и pH через ASIC3. Nat. Коммуна . 7: 10002. DOI: 10.1038 / ncomms10002
, Валлен, П., и Грилльнер, С. (2016). Ресничные нейроны, выстилающие центральный канал, воспринимают движение жидкости и pH через ASIC3. Nat. Коммуна . 7: 10002. DOI: 10.1038 / ncomms10002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хименес, А. Дж., Томе, М., Паес, П., Вагнер, К., Родригес, С., Fernández-Llebrez, P., et al. (2001). Запрограммированная денудация эпендимы предшествует врожденной гидроцефалии у мутантных мышей hyh. J. Neuropathol. Exp. Neurol . 60, 1105–1119. DOI: 10.1093 / jnen / 60.11.1105
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Капли П., Нацидис П., Лейте Д. Дж., Фурсман М., Джеффри Н., Рахман И. А. и др. (2021 г.). Отсутствие поддержки Deuterostomia побуждает к переосмыслению первой Bilateria. Sci. Adv .7: eabe2741. DOI: 10.1126 / sciadv.abe2741
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кавасима, Т., Кавашима, С., Танака, К., Мураи, М. , Йонеда, М., Патнам, Н. Х. и др. (2009). Перетасовка доменов и эволюция позвоночных. Genome Res . 19, 1393–1403. DOI: 10.1101 / gr.087072.108
, Йонеда, М., Патнам, Н. Х. и др. (2009). Перетасовка доменов и эволюция позвоночных. Genome Res . 19, 1393–1403. DOI: 10.1101 / gr.087072.108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Колмер, В. (1921). Das Sagittalorgan der wirbeltiere. Zeitschrift Anat.Entwicklungsgesch . 60, 652–717.
Google Scholar
Крамер-Цукер А. Г., Олале Ф., Хейкрафт К. Дж., Йодер Б. К., Шир А. Ф. и Драммонд И. А. (2005). Управляемый ресничками поток жидкости в пронефросе, мозге и пузырьке Купфера рыбок данио необходим для нормального органогенеза. Развитие 132, 1907–1921. DOI: 10.1242 / dev.01772
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лакалли, Т. К. (2000). Морфология клеток в нервном канатике амфиоксуса может отражать динамику клеточной дифференцировки. Внутр. J. Dev. Биол. 44, 903–906.
Google Scholar
Ли, К., Тан, Дж., Моррис, М. Б., Рицзоти, К., Хьюз, Дж., Чеа, П. С. и др. (2012). Врожденная гидроцефалия и аномальное развитие субкомиссуральных органов у трансгенных мышей Sox3. PLoS One 7: e29041. DOI: 10.1371 / journal.pone.0029041
С. и др. (2012). Врожденная гидроцефалия и аномальное развитие субкомиссуральных органов у трансгенных мышей Sox3. PLoS One 7: e29041. DOI: 10.1371 / journal.pone.0029041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lichtenfeld, J., Viehweg, J., Schutzenmeister, J., and Naumann, W. W. (1999).Вещество Рейсснера выражается в виде переходного рисунка в пластине дна позвоночных. Анат. Эмбриол. (Берл). 200, 161–174. DOI: 10.1007 / s0042
270
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоу, К. Дж., Кларк, Д. Н., Медейрос, Д. М., Рохсар, Д. С., и Герхарт, Дж. (2015). Контекст deuterostome происхождения хордовых. Природа 520, 456–465. DOI: 10.1038 / природа14434
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лу, Х., Шагирова А., Гогги Дж. Л., Йео Х. Л. и Рой С. (2020). Индуцированная волокном Рейсснера передача сигналов уротензина от нейронов, контактирующих с спинномозговой жидкостью, предотвращает сколиоз позвоночника. Biol. Откройте 9: bio052027. DOI: 10.1242 / bio.052027
Biol. Откройте 9: bio052027. DOI: 10.1242 / bio.052027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Машанов В. С., Зуева О. Р., Хайнцеллер Т., Ашауэр Б., Науманн В. В., Грондона Дж. М. и др. (2009). Центральная нервная система морских огурцов (Echinodermata: holothuroidea) показывает положительное иммуноокрашивание на хордовый глиальный секрет. Перед. Zool. 6:11. DOI: 10.1186 / 1742-9994-6-11
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мацумото Ю., Ямагути Ю., Хамачи М., Нономура К., Мурамацу Ю., Йошида Х. и др. (2020). Апоптоз участвует в поддержании характера среднего мозга и верхней пластинки промежуточного мозга после закрытия нервной трубки. Dev. Биол. 2020, 101–109. DOI: 10.1016 / j.ydbio.2020.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Майниэль, А., Meiniel, R., Didier, R., Creveaux, I., Gobron, S., Monnerie, H., et al. (1996). Подкомиссуральный орган и комплекс волокон Рейсснера. Загадка центральной нервной системы? Прог. Histochem. Cytochem. 30, 1–66. DOI: 10.1016 / s0079-6336 (96) 80015-5
Загадка центральной нервной системы? Прог. Histochem. Cytochem. 30, 1–66. DOI: 10.1016 / s0079-6336 (96) 80015-5
CrossRef Полный текст | Google Scholar
Мейниэль, О., Мейниэль, А. (2007). Сложная многодоменная организация белка SCO-спондина высоко консервативна у млекопитающих. Brain Res. Ред. 53, 321–327.DOI: 10.1016 / j.brainresrev.2006.09.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Meiniel, O., Meiniel, R., Lalloué, F., Didier, R., Jauberteau, M.O., Meiniel, A., et al. (2008). Удлинение гигантского белка: когда, как и почему? J. Mol. Evol. 66, 1–10. DOI: 10.1007 / s00239-007-9055-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Муньос, Р. И., Кане, Т., Эррера, Х., Родригес, С., Герра, М. М., Вио, К., и другие. (2019). Подкомиссуральный орган и волокно Рейсснера: возвращение к старым друзьям. Cell Tissue Res. 375, 507–529. DOI: 10.1007 / s00441-018-2917-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Николлс, Г. Э. (1913). Строение и развитие клетчатки Рейсснера и подкомиссурального органа. Q. J. Microsc. Sci. 58 (Часть 1), 1–116.
Э. (1913). Строение и развитие клетчатки Рейсснера и подкомиссурального органа. Q. J. Microsc. Sci. 58 (Часть 1), 1–116.
Google Scholar
Nieuwenhuys, R. (1988). «Amphioxus», в Центральная нервная система позвоночных, , ред.Ньювенхейс, Х. Дж. Тен Донкелаар и К. Николсон (Берлин: Springer), 365–396.
Google Scholar
Обермюллер-Вилен, Х. и Олссон, Р. (1974). Окончание волокна Райсснера в некоторых нижних хордовых. Acta Zool. 55, 71–79.
Google Scholar
Olsson, R. (1972). Волокно Рейсснера у личинок головастиков асцидии. Acta Zool. Stockh . 53, 17–21.
Google Scholar
Олссон Р. и Вингстранд К. Г. (1954).Волокно Рейсснера и инфундибулярный орган в Amphioxus. Univ. Берген Арбок (Publ. Biol. Stat.) 14, 1–14.
Google Scholar
Олссон Р., Юлис Р. и Родригес Э. М. (1994). Воронкообразный орган ланцетника (Branchiostoma lanceolatum Acrania): иммуноцитохимическое исследование.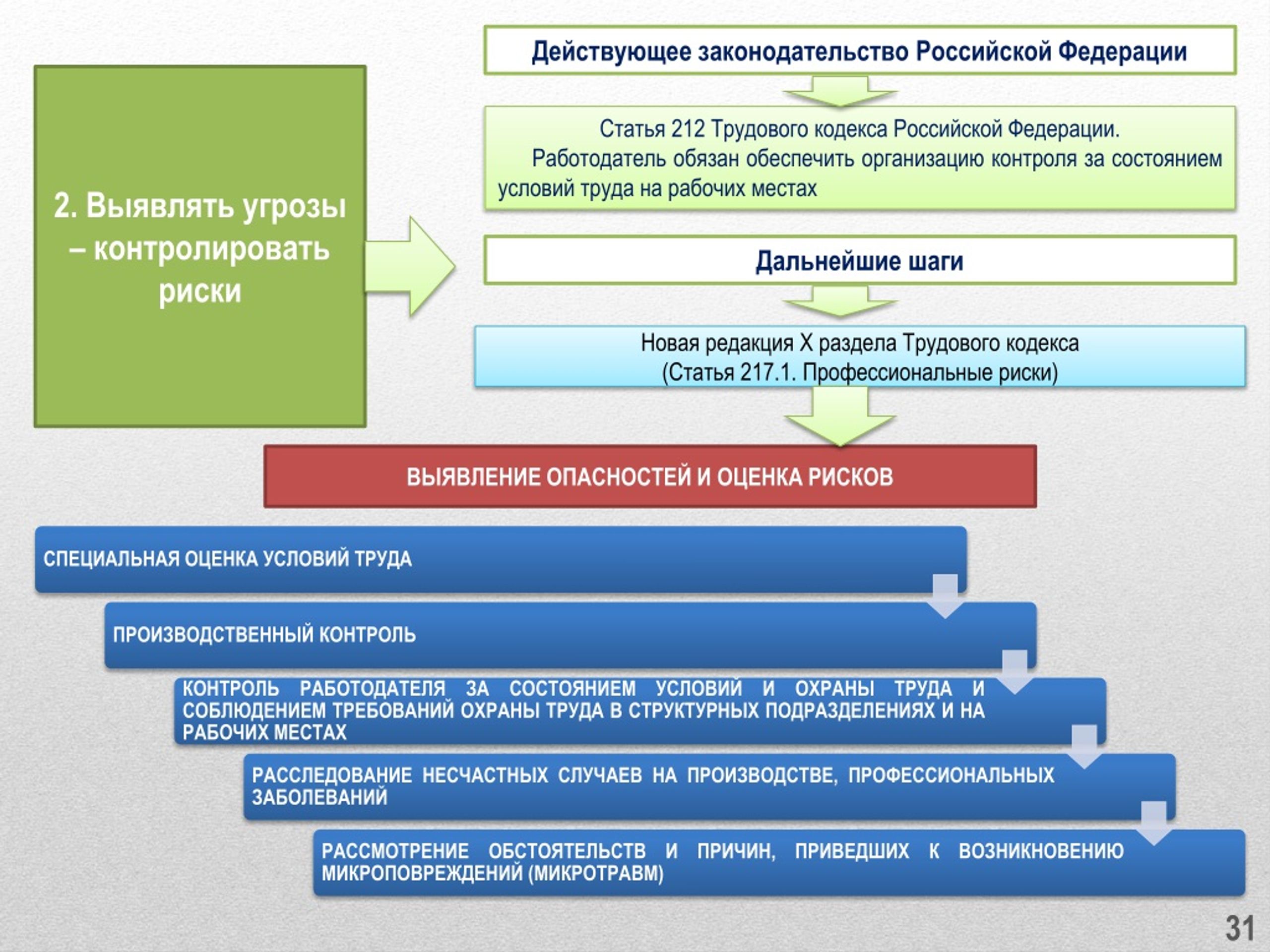 Cell Tiss. Res . 277, 107–114.
Cell Tiss. Res . 277, 107–114.
Google Scholar
Ортс-Дель’Иммаджин, А., Канто-Белариф, Ю., Тувенен, О., Руссель, Дж., Баскаран, А., Ланги, Д., и другие. (2020). Сенсорным нейронам, контактирующим со спинномозговой жидкостью, требуется волокно Рейсснера для обнаружения искривления позвоночника in vivo. Curr. Биол. 30, 827.e4–839.e4. DOI: 10.1016 / j.cub.2019.12.071
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оверхолзер М. Д., Уитли Дж. Р., О’Делл Б. Л. и Хоган А. Г. (1954). Желудочковая система головного мозга крыс с гидроцефалией вырабатывается дефицитом витамина B12 или фолиевой кислоты в рационе матери. Анат.Рек. 120, 917–933.
Google Scholar
Пани, А. М., Мулларки, Э. Э., Аронович, Дж., Ассимакопулос, С., Гроув, Э. А., и Лоу, К. Дж. (2012). Древние deuterostome происхождения сигнальных центров мозга позвоночных. Природа 483, 289–294. DOI: 10.1038 / nature10838
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес-Фигарес, Х. М., Хименес, А. Х. и Родригес, Э. М. (2001). Подкомиссуральный орган, циркуляция спинномозговой жидкости и гидроцефалия. Microsc. Res. Tech. 52, 591–607. DOI: 10.1002 / 1097-0029 (20010301) 52: 5 <591 :: aid-jemt1043 <3.0.co; 2-7
М., Хименес, А. Х. и Родригес, Э. М. (2001). Подкомиссуральный орган, циркуляция спинномозговой жидкости и гидроцефалия. Microsc. Res. Tech. 52, 591–607. DOI: 10.1002 / 1097-0029 (20010301) 52: 5 <591 :: aid-jemt1043 <3.0.co; 2-7
CrossRef Полный текст | Google Scholar
Рамос, К., Фернандес-Ллебрес, П., Бах, А., Роберт, Б., и Сориано, Э. (2004). Нарушение Msx1 приводит к дефектам промежуточного мозга и гидроцефалии. Dev. Дин. 230, 446–460. DOI: 10.1002 / dvdy.20070
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рейсснер, Э. (1860). Beiträge zur Kenntnis vom Bau des Rückenmarkes von Petromyzon fluviatilis L. Arch. Анат. Physiol. Wiss Med. (Лейпциг). 77, 545–588.
Google Scholar
Рихтер, Х. Г., Томе, М. М., Юлис, К. Р., Вио, К. Дж., Хименес, А. Х., Перес-Фигарес, Дж. М. и др. (2004). Транскрипция SCO-спондина в субкомиссуральном органе: данные о понижающей регуляции, опосредованной серотонином. Brain Res. Мол. Brain Res. 129, 151–162. DOI: 10.1016 / j.molbrainres.2004.07.003
Brain Res. Мол. Brain Res. 129, 151–162. DOI: 10.1016 / j.molbrainres.2004.07.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Родригес, Э.М., Окше, А., Хайн, С., Юлис, К. Р. (1992). Клеточная биология подкомиссурального органа. Внутр. Rev. Cytol. 135, 39–121. DOI: 10.1016 / s0074-7696 (08) 62038-0
CrossRef Полный текст | Google Scholar
Родригес, Э. М., Родригес, С., и Хайн, С. (1998). Подкомиссуральный орган. Microsc. Res. Тех . 41, 98–123. DOI: 10.1002 / (SICI) 1097-0029 (19980415) 41: 2 <98 :: AID-JEMT2 <3.0.CO; 2-M
CrossRef Полный текст | Google Scholar
Роуз, К.Д., Помпили, Д., Хенке, К., Ван Геннип, Дж. Л. М., Мейер-Майнер, А., Рана, Р. и др. (2020). Дефекты SCO-Spondin и нейровоспаление являются консервативными механизмами, вызывающими деформацию позвоночника, в генетических моделях идиопатического сколиоза. Curr. Биол. 30, 2363.e6–2373.e6. DOI: 10.1016 / j.cub.2020. 04.020
04.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сасай, Н., Кадоя, М., и Онг Ли Чен, А. (2021 г.). Нейронная индукция: исторические взгляды и применение к плюрипотентным стволовым клеткам. Dev. Разница в росте. 63, 26–37. DOI: 10.1111 / dgd.12703
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стридтер Г. Ф. и Норткатт Р. Г. (2020). Мозги сквозь время. Естественная история позвоночных. Лондон: Издательство Оксфордского университета.
Google Scholar
Troutwine, B.R., Gontarz, P., Konjikusic, M.J., Minowa, R., Monstad-Rios, A., Sepich, D. S., et al. (2020). Волокно Рейсснера очень динамично in vivo и контролирует морфогенез позвоночника. Curr. Биол . 30, 2353.e3–2362.e3. DOI: 10.1016 / j.cub.2020.04.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вио К., Родригес С., Наваррете Э. Х., Перес-Фигарес Х. М., Хименес А. Х. и Родригес Э. М. (2000). Гидроцефалия, вызванная иммунологической блокадой комплекса субкомиссурального органа с волокном Рейсснера (RF) в результате передачи от матери анти-RF антител. Exp. Brain Res. 135, 41–52. DOI: 10.1007 / s002210000474
Гидроцефалия, вызванная иммунологической блокадой комплекса субкомиссурального органа с волокном Рейсснера (RF) в результате передачи от матери анти-RF антител. Exp. Brain Res. 135, 41–52. DOI: 10.1007 / s002210000474
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вио, К., Родригес, С., Юлис, К. Р., Оливер, К., и Родригес, Э. М. (2008). Субкомиссуральный орган крысы секретирует гликопротеины клетчатки Рейсснера и растворимые в спинномозговой жидкости белки, достигающие внутреннего и внешнего отсеков спинномозговой жидкости. Цереброспинальная жидкость Res . 5: 3. DOI: 10.1186 / 1743-8454-5-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вагнер К., Батис Л. Ф., Родригес С., Хименес А. Х., Паес П., Томе М. и др. (2003). Клеточные механизмы, участвующие в стенозе и облитерации церебрального водопровода у мышей с мутантами hyh, у которых развивается врожденная гидроцефалия. J. Neuropathol. Exp. Neurol. 62, 1019–1040. DOI: 10.1093 / jnen / 62.10.1019
DOI: 10.1093 / jnen / 62.10.1019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ян С., Емельянов А., Ю М. С., Син М., Корж В. (2021). Верблюд регулирует развитие желудочковой системы головного мозга. Cell Tissue Res. 383, 835–852. DOI: 10.1007 / s00441-020-03270-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Специфические для опухолей амплитудно-модулированные радиочастотные электромагнитные поля вызывают дифференцировку гепатоцеллюлярной карциномы посредством нацеливания на Cav3.Напряжениезависимые кальциевые каналы 2T-типа и приток Ca2 +
https://doi.org/10.1016/j.ebiom.2019.05.034Получить права и контентАннотация
Предпосылки
Администрирование амплитудно-модулированных электромагнитных радиочастот 27 · 12 МГц полей (AM RF EMF) с помощью аппликатора в форме ложки, помещенного на язык пациента, – это недавно одобренный метод лечения запущенной гепатоцеллюлярной карциномы (HCC). Механизм действия опухолеспецифической AM RF EMF в значительной степени неизвестен.
Механизм действия опухолеспецифической AM RF EMF в значительной степени неизвестен.
Методы
Были выполнены дозиметрические анализы всего тела и органов человека.Мышей, несущих ксенотрансплантаты HCC человека, подвергали воздействию AM RF EMF с использованием системы воздействия AM RF EMF на мелких животных, воспроизводящей дозиметрию человека и время воздействия. Мы провели гистологический анализ опухолей после воздействия AM RF EMF. Используя агностический геномный подход, мы охарактеризовали механизм действия AM RF EMF.
Результаты
Внутрибукальное введение приводит к системной доставке атермальной AM RF EMF с головы до пят на уровнях ниже, чем те, которые генерируются сотовыми телефонами, которые держат близко к телу.Уменьшение опухоли происходит в результате дифференцировки клеток HCC в покоящиеся клетки с морфологией веретена. AM RF EMF нацеленные антипролиферативные эффекты и эффекты ингибирования раковых стволовых клеток опосредуются притоком Ca 2+ через Ca v 3 · 2T-типа потенциалзависимые кальциевые каналы (CACNA1H), что приводит к увеличению внутриклеточной концентрации кальция только в клетках HCC .
Интерпретация
AM RF EMF, вводимая через ротовую полость, представляет собой системную терапию, которая избирательно блокирует рост клеток HCC.AM RF EMF выраженный ингибирующий эффект на раковые стволовые клетки может объяснить исключительно длительные реакции, наблюдаемые у нескольких пациентов с запущенной стадией HCC.
Fund
Исследование, представленное в этой публикации, было поддержано грантом поддержки онкологического центра Национального института рака № P30CA012197, выданным Баптистскому комплексному онкологическому центру Уэйк Форест (BP), и фондами Профессорского фонда Чарльза Л. Спурра (BP). . DWG поддерживается R01 AA016852 и P50 AA026117.
Ключевые слова
Распространенная гепатоцеллюлярная карцинома
Т-образные потенциалозависимые кальциевые каналы
Приток кальция
Ca v 3 · 2
CACNA1H
Амплитудно-модулированные поля
RF RFРекомендуемые статьиЦитирующие статьи (0)
© 2019 Авторы. Опубликовано Elsevier B.V.
Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирующие статьи
Обновление 2020 г. рекомендаций WSES по ведению острого дивертикулита толстой кишки в условиях неотложной помощи | Всемирный журнал неотложной хирургии
Вайцман А.В., Нгуен Г.К. Дивертикулярная болезнь: эпидемиология и лечение. Можно J Гастроэнтерол. 2011; 25: 385–9.
PubMed PubMed Central Статья Google ученый
Schoetz DJ. Дивертикулярная болезнь толстой кишки: проблема вековой давности. Dis Colon Rectum. 1999; 42: 703–9.
PubMed Статья PubMed Central Google ученый
Shahedi K, Fuller G, Bolus R, Cohen E, Vu M, Shah R, et al.Долгосрочный риск острого дивертикулита у пациентов с случайным дивертикулезом, обнаруженный во время колоноскопии. Clin Gastroenterol Hepatol. 2013; 11: 1609–13.
PubMed PubMed Central Статья Google ученый

Коллинз Д., Уинтер, округ Колумбия. Современные концепции дивертикулярной болезни. J Clin Gastroenterol. 2015; 49: 358–69.
PubMed Статья PubMed Central Google ученый
Warner E, Crighton EJ, Moineddin R, Mamdani M, Upshur R. Четырнадцатилетнее исследование госпитализаций по поводу дивертикулярной болезни в Онтарио. Можно J Гастроэнтерол. 2007; 21: 97–9.
PubMed PubMed Central Статья Google ученый
Jamal Talabani A, Lydersen S, Endreseth BH, Edna TH. Значительное увеличение частоты госпитализаций и заболеваемости острым дивертикулитом толстой кишки. Int J Color Dis. 2014; 29: 937–45.
Артикул Google ученый
Сартелли М., Катена Ф., Ансалони Л., Кокколини Ф., Гриффитс Э.А., Абу-Зидан Ф.М. и др. Рекомендации WSES по ведению острого дивертикулита левой толстой кишки в условиях неотложной помощи. Мир J Emerg Surg. 2016; 11:37.
Мир J Emerg Surg. 2016; 11:37.
PubMed PubMed Central Статья Google ученый
Guyatt GH, Oxman AD, Vist GE, Kunz R, Falck-Ytter Y, Alonso-Coello P, et al. ОЦЕНКА: формирующийся консенсус по оценке качества доказательств и силы рекомендаций.BMJ. 2008; 336: 924–6.
PubMed PubMed Central Статья Google ученый
Brozek JL, Akl EA, Jaeschke R, Lang DM, Bossuyt P, Glasziou P, et al. Оценка качества доказательств и силы рекомендаций в руководствах по клинической практике: часть 2 из 3. Подход GRADE к оценке качества доказательств о диагностических тестах и стратегиях. Аллергия. 2009. 64: 1109–16.
CAS PubMed Статья PubMed Central Google ученый
Хинчи Э.Дж., Шаал PH, Ричардс МБ. Лечение перфоративной дивертикулярной болезни толстой кишки. Adv Surg. 1978; 12: 85–109.
CAS PubMed PubMed Central Google ученый
Neff CC, van Sonnenberg E. CT дивертикулита. Диагностика и лечение. Radiol Clin N Am. 1989; 27: 743–52.
CAS PubMed PubMed Central Google ученый
Амброзетти П., Беккер С., Терьер Ф. Дивертикулит толстой кишки: влияние визуализации на хирургическое лечение – проспективное исследование 542 пациентов. Eur Radiol. 2002; 12: 1145–9.
CAS PubMed Статья PubMed Central Google ученый
Kaiser AM, Jiang JK, Lake JP, Ault G, Artinyan A, Gonzalez-Ruiz C, et al. Ведение осложненного дивертикулита и роль компьютерной томографии. Am J Gastroenterol. 2005; 100: 910–7.
PubMed Статья PubMed Central Google ученый
Mora Lopez L, Serra Pla S, Serra-Aracil X, Ballesteros E, Navarro S. Применение модифицированной классификации Неффа к пациентам с неосложненным дивертикулитом. Цвет Dis. 2013; 15: 1442–7.
Применение модифицированной классификации Неффа к пациентам с неосложненным дивертикулитом. Цвет Dis. 2013; 15: 1442–7.
CAS Статья Google ученый
Саллинен В.Дж., Леппяниеми АК, Ментула П.Дж. Стадия острого дивертикулита на основании клинических, рентгенологических и физиологических параметров.J Trauma Acute Care Surg. 2015; 78: 543–51.
PubMed Статья PubMed Central Google ученый
Toorenvliet BR, Bakker RF, Breslau PJ, Merkus JW, Hamming JF. Дивертикулит толстой кишки: проспективный анализ диагностической точности и принятия клинических решений. Цвет Dis. 2010; 12: 179–86.
CAS Статья Google ученый
Ламерис В., ван Ранден А., ван Гулик Т.М., Буш О.Р., Винкельхаген Дж., Боссайт П.М. и др.Правило клинического решения для установления диагноза острого дивертикулита в отделении неотложной помощи. Dis Colon Rectum. 2010; 53: 896–904.
Dis Colon Rectum. 2010; 53: 896–904.
PubMed Статья PubMed Central Google ученый
Andeweg CS, Knobben L, Hendriks JC, Bleichrodt RP, van Goor H. Как диагностировать острый левосторонний дивертикулит толстой кишки: предложение по системе клинической оценки. Ann Surg. 2011; 253: 940–6.
PubMed Статья PubMed Central Google ученый
Кечагиас А., Раутио Т., Кечагиас Г., Мякеля Дж. Роль С-реактивного белка в прогнозировании клинической тяжести острого дивертикулита. Am Surg. 2014; 80: 391–5.
PubMed PubMed Central Google ученый
Ван де Валл Б.Дж., Драайзма В.А., ван дер Кайдж Р.Т., Констен Э.С., Визер М.Дж., Бродерс И.А. Значение маркеров воспаления и температуры тела при остром дивертикулите. Цвет Dis. 2013; 15: 621–6.
Артикул Google ученый
Mäkelä JT, Klintrup K, Takala H, Rautio T. Роль C-реактивного белка в прогнозировании тяжести острого дивертикулита в отделении неотложной помощи. Сканд Дж Гастроэнтерол. 2015; 50: 536–41.
PubMed Статья CAS PubMed Central Google ученый
Kechagias A, Sofianidis A, Zografos G, Leandros E, Alexakis N, Dervenis C. Индекс C-реактивного белка прогнозирует усиление тяжести острого дивертикулита сигмовидной кишки.Ther Clin Risk Manag. 2018; 14: 1847–53.
CAS PubMed PubMed Central Статья Google ученый
Gewurz H, Mold C, Siegel J, Fiedel B. C-реактивный белок и реакция острой фазы. Adv Intern Med. 1982; 27: 345–72.
CAS PubMed PubMed Central Google ученый
Ламерис В., ван Ранден А., Бипат С., Боссайт П.М., Бурмеестер М.А., Стокер Дж.Ступенчатая компрессионная ультрасонография и компьютерная томография при остром дивертикулите толстой кишки: метаанализ точности теста. Eur Radiol. 2008. 18: 2498–511.
PubMed Статья PubMed Central Google ученый
Лильегрен Г., Чабок А., Викбом М., Смед К., Нильссон К. Острый дивертикулит толстой кишки: систематический обзор диагностической точности. Цвет Dis. 2007; 9: 480–8.
CAS Статья Google ученый
Амброзетти П., Дженни А., Беккер С., Терьер Т.Ф., Морель П. Острый дивертикулит левой толстой кишки – сравнение результатов компьютерной томографии и водорастворимой контрастной клизмы: проспективная оценка 420 пациентов. Dis Colon Rectum. 2000; 43: 1363–7.
CAS PubMed Статья PubMed Central Google ученый
Сартелли М., Мур Ф.А., Ансалони Л., Ди Саверио С., Кокколини Ф., Гриффитс Э.А. и др. Предложение по классификации острого дивертикулита левой кишки на основе компьютерной томографии.Мир J Emerg Surg. 2015; 10: 3.
PubMed PubMed Central Статья Google ученый
Mazzei MA, Cioffi Squitieri N, Guerrini S, Stabile Ianora AA, Cagini L., Macarini L., et al. Сигмовидный дивертикулит: данные США. Crit Ultrasound J. 2013; 5 (Suppl 1): S1–5.
Артикул Google ученый
Puylaert JB. УЗИ дивертикулита толстой кишки.Dig Dis. 2012; 30: 56–9.
PubMed Статья PubMed Central Google ученый
Andeweg CS, Wegdam JA, Groenewoud J, van der Wilt GJ, van Goor H, Bleichrodt RP. На пути к научно обоснованному подходу к диагностике дивертикулита. Сканд Дж Гастроэнтерол. 2014; 49: 775–84.
PubMed Статья PubMed Central Google ученый
Ламерис В., ван Ранден А., ван Эс Х.В., ван Хизвейк Дж. П., ван Рамсхорст Б., Баума WH и др.Исследовательская группа OPTIMA: стратегии визуализации для выявления неотложных состояний у пациентов с острой болью в животе: исследование диагностической точности. BMJ. 2009; 338: b2431.
PubMed PubMed Central Статья Google ученый
Heverhagen JT, Sitter H, Zielke A, Klose KJ. Проспективная оценка значения магнитно-резонансной томографии при подозрении на острый дивертикулит сигмовидной кишки. Dis Colon Rectum. 2008; 51: 1810–5.
PubMed Статья PubMed Central Google ученый
Halpenny DF, McNeil G, Snow A, Geoghegan T, Torreggiani WC. Проспективная оценка значения магнитно-резонансной томографии при подозрении на острый дивертикулит сигмовидной кишки. Dis Colon Rectum. 2009; 52: 1030–1.
PubMed Статья PubMed Central Google ученый
Hwang SS, Cannom RR, Abbas MA, Etzioni D. Дивертикулит у пациентов с трансплантатами и пациентов, получающих хроническую кортикостероидную терапию: систематический обзор. Dis Colon Rectum.2010; 53: 1699–707.
PubMed Статья PubMed Central Google ученый
Далла Валле Р., Капокасале Е., Маццони М.П., Буси Н., Беноцци Л., Сивелли Р. и др. Острый дивертикулит с перфорацией толстой кишки при трансплантации почки. Transplant Proc. 2005; 37: 2507–10.
CAS PubMed Статья PubMed Central Google ученый
Qasabian RA, Meagher AP, Lee R, Dore GJ, Keogh A.Тяжелый дивертикулит после трансплантации сердца, легких и сердце-легкое. J Пересадка сердца и легких. 2004; 23: 845–9.
PubMed Статья PubMed Central Google ученый
Lederman ED, Conti DJ, Lempert N, Singh TP, Lee EC. Осложненный дивертикулит после трансплантации почки. Dis Colon Rectum. 1998. 41: 613–8.
CAS PubMed Статья PubMed Central Google ученый
Bordeianou L, Hodin R. Споры в хирургическом лечении дивертикулита сигмовидной кишки. J Gastrointest Surg. 2007; 11: 542–8.
PubMed PubMed Central Статья Google ученый
Биондо С., Тренти Л., Эльвира Дж., Голда Т., Крейслер Э. Исходы дивертикулита толстой кишки по причине иммуносупрессии. Am J Surg. 2016; 21: 384–90.
Артикул Google ученый
Sartelli M, Chichom-Mefire A, Labricciosa FM, Hardcastle T, Abu-Zidan FM, Adesunkanmi AK, et al. Управление внутрибрюшными инфекциями с глобальной точки зрения: рекомендации WSES 2017 по ведению внутрибрюшных инфекций. Мир J Emerg Surg. 2017; 12:29. Ошибка в: World J Emerg Surg. 2017; 12:36.
Шабанзаде Д.М., Вилле-Йоргенсен П. Антибиотики при неосложненном дивертикулите. Кокрановская база данных Syst Rev.2012; 11: CD009092.
PubMed PubMed Central Google ученый
Чабок А., Полман Л., Хьерн Ф., Хаапаниеми С., Смед К., Исследовательская группа AVOD. Рандомизированное клиническое исследование антибиотиков при остром неосложненном дивертикулите. Br J Surg. 2012; 99: 532–9.
CAS PubMed Статья PubMed Central Google ученый
Mali JP, Mentula PJ, Leppäniemi AK, Sallinen VJ. Симптоматическое лечение неосложненного острого дивертикулита: проспективное когортное исследование. Dis Colon Rectum. 2016; 59: 529–34.
PubMed Статья PubMed Central Google ученый
Daniels L, Ünlü Ç, de Korte N, van Dieren S, Stockmann HB, Vrouenraets BC, et al. Рандомизированное клиническое испытание наблюдательного лечения в сравнении с лечением антибиотиками при первом эпизоде неосложненного острого дивертикулита, подтвержденного КТ. Br J Surg. 2017; 104: 52–61.
CAS PubMed Статья PubMed Central Google ученый
van Dijk ST, Daniels L, Ünlü Ç, de Korte N, van Dieren S, Stockmann HB, et al. Долгосрочные последствия отказа от приема антибиотиков при неосложненном остром дивертикулите. Am J Gastroenterol. 2018; 113: 1045–52.
PubMed Статья CAS PubMed Central Google ученый
Sartelli M, Viale P, Catena F, Ansaloni L, Moore E, Malangoni M, et al. Рекомендации WSES 2013 г. по ведению внутрибрюшных инфекций. Мир J Emerg Surg.2013; 8: 3.
PubMed PubMed Central Статья Google ученый
Риджуэй П.Ф., Латиф А., Шаббир Дж., Офриокума Ф., Херли М.Дж., Эвой Д. и др. Рандомизированное контролируемое исследование пероральной и внутривенной терапии клинически диагностированного острого неосложненного дивертикулита. Цвет Dis. 2009; 11: 941–6.
CAS Статья Google ученый
Fozard JB, Armitage NC, Schofield JB, Jones OM.Ассоциация колопроктологов Великобритании и Ирландии. Изложение позиции ACPGBI по поводу плановой резекции дивертикулита. Цвет Dis. 2011; 13: 1–11.
Артикул Google ученый
Etzioni DA, Chiu VY, Cannom RR, Burchette RJ, Haigh PI, Abbas MA. Амбулаторное лечение острого дивертикулита: показатели и предикторы неэффективности. Dis Colon Rectum. 2010; 53: 861–5.
PubMed Статья PubMed Central Google ученый
Джексон Дж. Д., Хаммонд Т. Систематический обзор: амбулаторное ведение острого неосложненного дивертикулита. Int J Color Dis. 2014; 29: 775–81.
CAS Статья Google ученый
Родригес-Серилло М., Поза-Монторо А, Фернандес-Диас Е., Матесанц-Давид М., Инурриета, РА. Лечение пожилых пациентов с неосложненным дивертикулитом, даже при сопутствующей патологии, в домашних условиях. Eur J Intern Med. 2013; 24: 430–2.
PubMed Статья PubMed Central Google ученый
Biondo S, Golda T, Kreisler E, Espin E, Vallribera F, Oteiza F, et al. Амбулаторное лечение неосложненного дивертикулита по сравнению с госпитализацией: проспективное многоцентровое рандомизированное клиническое исследование (исследование DIVER). Ann Surg. 2014; 259: 38–44.
PubMed Статья PubMed Central Google ученый
Cirocchi R, Randolph JJ, Binda GA, Gioia S, Henry BM, Tomaszewski KA, et al. Безопасно и эффективно ли амбулаторное ведение острого дивертикулита? Систематический обзор и метаанализ.Tech Coloproctol. 2019; 23: 87–100.
CAS PubMed Статья PubMed Central Google ученый
Balasubramanian I, Fleming C, Mohan HM, Schmidt K, Haglind E, Winter DC. Амбулаторное ведение дивертикулита легкой или неосложненной формы: систематический обзор. Dig Surg. 2017; 34: 151–60.
PubMed Статья PubMed Central Google ученый
Bolkenstein HE, van Dijk ST, Consten ECJ, Heggelman BGF, Hoeks CMA, Broeders IAMJ, et al. Консервативное лечение больных дивертикулитом периколичевым экстрапросветным воздухом и роль антибиотикотерапии. J Gastrointest Surg. 2019; 23: 2269–76.
CAS PubMed PubMed Central Статья Google ученый
Andersen JC, Bundgaard L, Elbrønd H, Laurberg S, Walker LR, Støvring J, et al. Датские национальные рекомендации по лечению дивертикулярной болезни.Дэн Мед Дж. 2012; 59: C4453.
PubMed PubMed Central Google ученый
Грегерсен Р., Мортенсен Л.К., Бурхарт Дж., Поммергаард Х.С., Розенберг Дж. Лечение пациентов с острым дивертикулитом толстой кишки, осложненным формированием абсцесса: систематический обзор. Int J Surg. 2016; 35: 201–8.
PubMed Статья PubMed Central Google ученый
Ambrosetti P, Chautems R, Soravia C, Peiris-Waser N, Terrier F.Отдаленный исход мезоколенного и тазового дивертикулярных абсцессов левой ободочной кишки: проспективное исследование 73 случаев. Dis Colon Rectum. 2005. 48: 787–91.
PubMed Статья PubMed Central Google ученый
Брандт Д., Герваз П., Дурмиши Ю., Платон А., Морел П., Полетти П.А. Чрескожный дренаж под контролем КТ по сравнению с одной антибиотикотерапией при дивертикулите Хинчи II: исследование случай-контроль. Dis Colon Rectum. 2006; 49: 1533–8.
CAS PubMed Статья PubMed Central Google ученый
Сиверт Б., Тай Дж., Крускал Дж., Сосна Дж., Опелка Ф., Раптопулос В. и др. Влияние дренирования под контролем КТ в лечении дивертикулярных абсцессов: размер имеет значение. AJR. 2006; 186: 680–6.
PubMed Статья PubMed Central Google ученый
Singh B, May K, Coltart I, Moore NR, Cunningham C.Отдаленные результаты чрескожного дренирования дивертикулярного абсцесса. Ann R Coll Surg Engl. 2008; 90: 297–301.
CAS PubMed PubMed Central Статья Google ученый
Кумар Р. Р., Ким Дж. Т., Хаукоос Дж. С., Масиас Л. Х., Диксон М. Р., Стамос М. Дж. И др. Факторы, влияющие на успешное лечение внутрибрюшных абсцессов антибиотиками и необходимость чрескожного дренирования. Dis Colon Rectum. 2006; 49: 183–9.
PubMed Статья PubMed Central Google ученый
Elagili F, Stocchi L, Ozuner G, Kiran RP. Использование только антибиотиков вместо чрескожного дренирования в качестве начального лечения большого дивертикулярного абсцесса. Tech Coloproctol. 2015; 19: 97–103.
CAS PubMed Статья PubMed Central Google ученый
Tsai HL, Hsieh JS, Yu FJ, Wu DC, Chen FM, Huang CJ, et al. Перфорированный рак толстой кишки, проявляющийся внутрибрюшным абсцессом. Int J Color Dis. 2007; 22: 15–9.
Артикул Google ученый
Yeo ES, Ng KH, Eu KW. Перфорированный колоректальный рак: важный дифференциальный диагноз при всех предполагаемых дивертикулярных абсцессах. Ann Acad Med Singap. 2011; 40: 375–8.
PubMed PubMed Central Google ученый
de Vries HS, Boerma D, Timmer R, van Ramshorst B, Dieleman LA, van Westreenen HL. Обычная колоноскопия не требуется при неосложненном дивертикулите: систематический обзор. Surg Endosc. 2014; 28: 2039–47.
PubMed Статья PubMed Central Google ученый
Sharma PV, Eglinton T, Hider P, Frizelle F. Систематический обзор и метаанализ роли рутинной оценки толстой кишки после радиологически подтвержденного острого дивертикулита. Ann Surg. 2014; 259: 263–72.
PubMed Статья PubMed Central Google ученый
Sallinen V, Mentula P, Leppäniemi A.Риск рака толстой кишки после острого дивертикулита, диагностированного с помощью компьютерной томографии: нужна ли рутинная колоноскопия? Surg Endosc. 2014; 28: 961–5.
PubMed Статья PubMed Central Google ученый
Фейнголд Д., Стил С.Р., Ли С., Кайзер А., Боуши Р., Буйе В.Д. и др. Параметры практики лечения дивертикулита сигмовидной кишки. Dis Colon Rectum. 2014; 57: 284–94.
PubMed Статья PubMed Central Google ученый
Дхармараджан С., Хант С.Р., Бирнбаум Э.Х., Флешман Дж. В., Матч М.Г. Эффективность консервативного лечения острого осложненного дивертикулита. Dis Colon Rectum. 2011; 54: 663–71.
PubMed Статья PubMed Central Google ученый
Саллинен В.Дж., Ментула П.Дж., Леппяниеми АК. Безоперационное лечение перфорированного дивертикулита с помощью экстрапросветного воздуха безопасно и эффективно у отдельных пациентов. Dis Colon Rectum. 2014; 57: 875–81.
PubMed Статья PubMed Central Google ученый
Росси Г.Л., Менц Р., Бертоне С., Охеа Кинтана Г., Бильбао С., Им В.М. и др. Лапароскопический лаваж брюшины при дивертикулите Хинчи III: насколько он эффективен и применим? Dis Colon Rectum. 2014; 57: 1384–90.
PubMed Статья PubMed Central Google ученый
Swank HA, Mulder IM, Hoofwijk AG, Nienhuijs SW, Lange JF, Bemelman WA.Голландское совместное исследование дивертикулярной болезни G. Ранний опыт лапароскопического лаважа при перфорированном дивертикулите. Br J Surg. 2013; 100: 704–10.
CAS PubMed Статья PubMed Central Google ученый
Angenete E, Thornell A, Burcharth J, Pommergaard HC, Skullman S, Bisgaard T, et al. Лапароскопический лаваж возможен и безопасен для лечения перфорированного дивертикулита с гнойным перитонитом: первые результаты рандомизированного контролируемого исследования DILALA.Ann Surg. 2016; 263: 117–22.
PubMed Статья PubMed Central Google ученый
Schultz JK, Yaqub S, Wallon C, Blecic L, Forsmo HM, Folkesson J, et al. Лапароскопический лаваж по сравнению с первичной резекцией при остром перфорированном дивертикулите: рандомизированное клиническое исследование SCANDIV. ДЖАМА. 2015; 314: 1364–75.
CAS PubMed Статья PubMed Central Google ученый
Vennix S, Musters GD, Mulder IM, Swank HA, Consten EC, Belgers EH, et al. Лапароскопический перитонеальный лаваж или сигмоидэктомия при перфорированном дивертикулите с гнойным перитонитом: многоцентровое рандомизированное открытое исследование в параллельных группах. Ланцет. 2015; 386 (10000): 1269–77.
PubMed Статья PubMed Central Google ученый
Ceresoli M, Coccolini F, Montori G, Catena F, Sartelli M, Ansaloni L. Сравнение лапароскопического лаважа и резекции при перфорированном дивертикулите с гнойным перитонитом: метаанализ рандомизированных контролируемых исследований.Мир J Emerg Surg. 2016; 11:42.
PubMed PubMed Central Статья Google ученый
Пенна М., Маркар С.Р., Маккензи Х., Хомпес Р., Каннингем С. Лапароскопический лаваж по сравнению с первичной резекцией при остром перфорированном дивертикулите: обзор и метаанализ. Ann Surg. 2018; 267: 252–8.
PubMed Статья PubMed Central Google ученый
Гэлбрейт Н., Картер Дж. В., Нетц У., Ян Д., Фрай Д. Э., Маккафферти М. и др. Лапароскопический лаваж в лечении перфорированного дивертикулита: современный метаанализ. J Gastrointest Surg. 2017; 21: 1491–9.
PubMed Статья PubMed Central Google ученый
Чирокки Р., Ди Саверио С., Вебер Д.Г., Табола Р., Абраха И., Рэндольф Дж. И др. Лапароскопический лаваж против хирургической резекции при остром дивертикулите с генерализованным перитонитом: систематический обзор и метаанализ.Tech Coloproctol. 2017; 21: 93–110.
CAS PubMed Статья PubMed Central Google ученый
Angenete E, Bock D, Rosenberg J, Haglind E. Лапароскопический лаваж превосходит резекцию толстой кишки при перфорированном гнойном дивертикулите – метаанализ. Int J Color Dis. 2017; 32: 163–9.
Артикул Google ученый
Shaikh FM, Stewart PM, Walsh SR, Davies RJ.Лапароскопический перитонеальный лаваж или хирургическая резекция при остром перфорированном дивертикулите сигмовидной кишки: систематический обзор и метаанализ. Int J Surg. 2017; 38: 130–7.
PubMed Статья PubMed Central Google ученый
Biffl WL, Moore FA, Moore EE. Возрастающая роль лапароскопического лаважа и дренажа. J Trauma Acute Care Surg. 2019; 86: 376.
PubMed Статья PubMed Central Google ученый
McCafferty MH, Roth L, Jorden J. Текущее лечение дивертикулита. Am Surg. 2008; 74: 1041–9.
PubMed PubMed Central Google ученый
Fleming FJ, Gillen P. Отмена процедуры Хартмана после острого дивертикулита: все ли рассчитано? Int J Color Dis. 2009; 24: 1219–25.
Артикул Google ученый
Hong MK, Tomlin AM, Hayes IP, Skandarajah AR.Частота оперативных вмешательств при остром дивертикулите: многоцентровое исследование в масштабе штата. ANZ J Surg. 2015; 85: 734–8.
PubMed Статья PubMed Central Google ученый
Ли Д., Бакстер Н.Н., Маклеод Р.С., Мойеддин Р., Уилтон А.С., Натенс А.Б. Развитие моделей практики лечения острого дивертикулита толстой кишки: популяционный анализ. Dis Colon Rectum. 2014; 57: 1397–405.
PubMed Статья PubMed Central Google ученый
Чандра V, Нельсон Х, Ларсон Д.Р., Харрингтон-младший. Влияние первичной резекции на исход пациентов с перфорированным дивертикулитом. Arch Surg. 2004; 139: 1221–4.
PubMed Статья PubMed Central Google ученый
Салем Л., Флум ДР. Первичный анастомоз или процедура Гартмана для пациентов с дивертикулярным перитонитом? Систематический обзор. Dis Colon Rectum. 2004; 47: 1953–64.
PubMed Статья PubMed Central Google ученый
Аббас С. Резекция и первичный анастомоз при остром осложненном дивертикулите, систематический обзор литературы. Int J Color Dis. 2007; 22: 351–7.
Артикул Google ученый
Чирокки Р., Трастулли С., Дезидерио Дж., Листорти С., Бозелли С., Паризи А. и др. Лечение дивертикулита III-IV стадии по Хинчи: систематический обзор и метаанализ. Int J Color Dis. 2013; 28: 447–57.
Артикул Google ученый
Ли Дж. М., Бай П., Чанг Дж., Эль-Хечи М., Конгкаевпайсан Н., Бонд А. и др. Сравнение процедуры Хартмана с первичным анастомозом с илеостомией с отклоняющейся петлей при остром дивертикулите: общенациональный анализ 2729 пациентов, перенесших экстренную операцию. J Am Coll Surg. 2019; 229: 48–55.
PubMed Статья PubMed Central Google ученый
Константинидес В.А., Хериот А., Ремзи Ф., Дарзи А., Сенапати А., Фазио В.В. и др. Оперативные стратегии дивертикулярного перитонита: анализ выбора между первичной резекцией и анастомозом по сравнению с процедурами Хартмана.Ann Surg. 2007. 245: 94–103.
PubMed PubMed Central Статья Google ученый
Оберкофлер К.Э., Рикенбахер А., Раптис Д.А., Леманн К., Виллигер П., Бухли С. и др. Многоцентровое рандомизированное клиническое исследование первичного анастомоза или процедуры Хартмана при перфорированном дивертикулите левой толстой кишки с гнойным или фекальным перитонитом. Ann Surg. 2012; 256: 819–26.
PubMed Статья PubMed Central Google ученый
Bridoux V, Regimbeau JM, Ouaissi M, Mathonnet M, Mauvais F, Houivet E, et al. Процедура Хартмана или первичный анастомоз при генерализованном перитоните вследствие перфоративного дивертикулита: проспективное многоцентровое рандомизированное исследование (DIVERTI). J Am Coll Surg. 2017; 225: 798–805.
PubMed Статья PubMed Central Google ученый
Lambrichts DPV, Vennix S, Musters GD, Mulder IM, Swank HA, Hoofwijk AGM, et al. Сравнение процедуры Хартмана с сигмоидэктомией с первичным анастомозом при перфорированном дивертикулите с гнойным или фекальным перитонитом (LADIES): многоцентровое рандомизированное открытое исследование превосходства в параллельных группах.Ланцет Гастроэнтерол Гепатол. 2019; 4: 599–610.
PubMed Статья PubMed Central Google ученый
Халим Х., Аскари А., Нанн Р., Холлингсхед Дж. Первичный анастомоз резекции по сравнению с процедурой Хартмана при дивертикулите Хинчи III и IV. Мир J Emerg Surg. 2019; 14:32.
PubMed PubMed Central Статья Google ученый
Vennix S, Boersema GS, Buskens CJ, Menon AG, Tanis PJ, Lange JF, et al.Неотложная лапароскопическая сигмоидэктомия по поводу перфоративного дивертикулита с генерализованным перитонитом: систематический обзор. Dig Surg. 2016; 33: 1–7.
PubMed Статья PubMed Central Google ученый
Weber DG, Bendinelli C, Balogh ZJ. Операция по борьбе с повреждениями при неотложных состояниях брюшной полости. Br J Surg. 2014; 101: e109–18.
CAS PubMed Статья PubMed Central Google ученый
Tartaglia D, Costa G, Camillò A, Castriconi M, Andreano M, Lanza M и др. Операция по устранению повреждений при перфорированном дивертикулите с диффузным перитонитом: спасает жизни и сокращает количество стом. Мир J Emerg Surg. 2019; 14:19.
PubMed PubMed Central Статья Google ученый
Kafka-Ritsch R, Birkfellner F, Perathoner A, Raab H, Nehoda H, Pratschke J, et al. Хирургия контроля повреждений с абдоминальным вакуумом и отсроченной реконструкцией кишечника у пациентов с перфорированным дивертикулитом. Hinchey III / IV.J Gastrointest Surg. 2012; 16: 1915–22.
PubMed Статья PubMed Central Google ученый
Perathoner A, Klaus A, Mühlmann G, Oberwalder M, Margreiter R, Kafka-Ritsch R. Контроль повреждений с помощью абдоминальной вакуумной терапии (VAC) для лечения перфорированного дивертикулита с распространенным генерализованным перитонитом – доказательство концепции. Int J Color Dis. 2010; 25: 767–74.
Артикул Google ученый
Ordóñez CA, Sánchez AI, Pineda JA, Badiel M, Mesa R, Cardona U, et al. Отложенный первичный анастомоз по сравнению с отвлечением у пациентов с тяжелым вторичным перитонитом, которым проводилась поэтапная лапаротомия. Мир J Surg. 2010; 34: 169–76.
PubMed PubMed Central Статья Google ученый
Зон М., Ага А., Хейтланд В., Гандлинг Ф., Штайнер П., Иесалниекс И. Стратегия контроля повреждений при лечении перфорированного дивертикулита с генерализованным перитонитом.Tech Coloproctol. 2016; 20: 577–83.
CAS PubMed Статья PubMed Central Google ученый
Сартелли М., Катена Ф., Абу-Зидан Ф.М., Ансалони Л., Биффл В.Л., Бурмеестер М.А. и др. Управление интраабдоминальными инфекциями: рекомендации консенсусной конференции WSES 2016. Мир J Emerg Surg. 2017; 12:22.
PubMed PubMed Central Статья Google ученый
Киркпатрик А. В., Кокколини Ф., Ансалони Л., Робертс Д. Д., Толонен М., Макки Дж. Л. и др. Закрытые или открытые после лапаротомии (COOL) после контроля источника для исследователей тяжелого осложненного внутрибрюшного сепсиса. Закрытое или открытое после лапаротомии в системе контроля источника при тяжелом осложненном внутрибрюшном сепсисе (исследование COOL): протокол рандомизированного контролируемого исследования. Мир J Emerg Surg. 2018; 13:26.
PubMed PubMed Central Статья Google ученый
Рафферти Дж., Шеллито П., Хайман Нью-Хэмпшир, Буйе, Вирджиния. Комитет по стандартам Американского общества хирургов толстой и прямой кишки. Параметры практики при дивертикулите сигмовидной кишки. Dis Colon Rectum. 2006; 49: 939–44.
PubMed Статья PubMed Central Google ученый
Холл Дж. Ф., Робертс П. Л., Риккарди Р., Рид Т., Шейри К., Уолд С. и др. Долгосрочное наблюдение после первого эпизода дивертикулита: каковы предикторы рецидива? Dis Colon Rectum.2011; 54: 283–8.
PubMed Статья PubMed Central Google ученый
Humes DJ, West J. Роль острого дивертикулита в развитии осложненного дивертикулярного заболевания толстой кишки и 1-летняя смертность после постановки диагноза в Великобритании: популяционная когорта. Кишечник. 2012; 61: 95–100.
PubMed Статья PubMed Central Google ученый
Салем Т.А., Моллой Р.Г., О’Дуайер П.Дж.Проспективное пятилетнее катамнестическое исследование пациентов с симптоматической неосложненной дивертикулярной болезнью. Dis Colon Rectum. 2007; 50: 1–5.
Артикул Google ученый
Эль-Сайед С., Рэдли С., Миттон Дж., Эвисон Ф., Уорд СТ. Риск повторного заболевания и хирургического вмешательства после госпитализации по поводу острого дивертикулита. Dis Colon Rectum. 2018; 61: 382–9.
PubMed Статья PubMed Central Google ученый
Regenbogen SE, Hardiman KM, Hendren S, Morris AM. Хирургия дивертикулита в 21 веке: систематический обзор. JAMA Surg. 2014; 149: 292–303.
PubMed Статья PubMed Central Google ученый
van de Wall BJM, Stam MAW, Draaisma WA, Stellato R, Bemelman WA, Boermeester MA, et al. Хирургия в сравнении с консервативным лечением рецидивирующего и продолжающегося левостороннего дивертикулита (исследование DIRECT): открытое многоцентровое рандомизированное контролируемое исследование.Ланцет Гастроэнтерол Гепатол. 2017; 2: 13–22.
PubMed Статья PubMed Central Google ученый
Klarenbeek BR, Samuels M, van der Wal MA, van der Peet DL, Meijerink WJ, Cuesta MA. Показания к плановой резекции сигмовидной кишки при дивертикулярной болезни. Ann Surg. 2010; 251: 670–4.
PubMed Статья PubMed Central Google ученый
Sawyer RG, Claridge JA, Nathens AB, Rotstein OD, Duane TM, Evans HL, et al. Испытание краткосрочного курса антимикробной терапии внутрибрюшной инфекции. NEJM. 2015; 372: 1996–2005.
PubMed Статья PubMed Central Google ученый
Chiu TC, Chou YH, Tiu CM, Chiou HJ, Wang HK, Lai YC, et al. Правосторонний дивертикулит толстой кишки: клинические особенности, сонографические проявления и лечение. J Med Ultrasound. 2017; 25: 33–9.
PubMed PubMed Central Статья Google ученый
Радхи Дж. М., Рамзи Дж. А., Бутросс-Тадросс О. Дивертикулярная болезнь правой ободочной кишки. BMC Res Notes. 2011; 4: 1–5.
Артикул Google ученый
Ha GW, Lee MR, Kim JH. Эффективность консервативного лечения у пациентов с дивертикулитом правой толстой кишки. ANZ J Surg. 2017; 87: 467–70.
PubMed Статья PubMed Central Google ученый
Little A, Culver A. Правосторонняя дивертикулярная перфорация сигмовидной кишки. West J Emerg Med. 2012; 13: 103–5.
PubMed PubMed Central Статья Google ученый
Espinosa J, Sharma R, Lucerna A, Stranges D. Медицинский доступ к дивертикулиту правой кишки с перфорацией. Case Rep Emerg Med. 2017; 2017: 2563218.
PubMed PubMed Central Google ученый
Kim SH, Byun CG, Cha JW, Choi SH, Kho YT, Seo DY. Сравнительное изучение клиники и лечения дивертикулита правой и левой толстой кишки. J Korean Soc Coloproctol. 2010; 26: 407–12.
PubMed PubMed Central Статья Google ученый
аль-Хилали MA, Razzaq HA, el-Salfiti JI, Abu-Zidan FM, al-Manee MS. Солитарный дивертикулит пищеварительной системы. Признание и менеджмент. Acta Chir Scand. 1989; 155: 475-8.
Kim JY, Park SG, Kang HJ, Lim YA, Pak KH, Yoo T и др. Проспективное рандомизированное клиническое исследование неосложненного правостороннего дивертикулита толстой кишки: антибиотики в сравнении с отсутствием антибиотиков. Int J Colorectal Dis. 2019; 34: 1413–20.
PubMed Статья PubMed Central Google ученый
Destek S, Gül VO. Эффективность консервативного подхода при дивертикулите правой кишки. Ulus Travma Acil Cerrahi Derg. 2019; 25: 396–402.
PubMed PubMed Central Google ученый
Chung BH, Ha GW, Lee MR, Kim JH. Ведение дивертикулита толстой кишки с учетом локализации и степени тяжести: сравнение правой и левой толстой кишки. Ann Coloproctol. 2016; 32: 228–33.
PubMed PubMed Central Статья Google ученый
Hildebrand P, Birth M, Bruch HP, Schwandner O.Хирургическое лечение правостороннего дивертикулита. Zentralbl Chir. 2005; 130: 123–7.
CAS PubMed Статья PubMed Central Google ученый
Можно ли восстановить точную клеточную линию с помощью регистраторов CRISPR?
Как вы увидите, рецензенты согласны с тем, что эта работа является ценным дополнением к быстрорастущей области регистраторов CRISPR и отслеживания происхождения. Каждый из них также вызывает озабоченность по поводу представленных результатов и их интерпретации.В частности, был достигнут четкий консенсус относительно соответствующего определения и требований к «точной» реконструкции родословной, обоснования используемых алгоритмов реконструкции дерева и методологий, используемых для анализа редактирования в целевых массивах дрозофилы. Рецензент 2 также имеет конкретные предложения по названию. Мы надеемся, что эти моменты будет легко оценить в отредактированной рукописи.
Благодарим рецензентов за конструктивные комментарии. Мы отредактировали рукопись, чтобы рассмотреть и прояснить поднятые вопросы, уделяя особое внимание оценке и интерпретации точности, а также уточнению нашей методологии.
Важные сообщения нашей статьи заключаются в следующем: всегда будет минимальный уровень точности, который должен быть достигнут, если реконструированные родственные отношения должны интерпретироваться уверенно; что это будет зависеть от проблемы; и что это требование необходимо учитывать при разработке эксперимента по клонированию.
Наличие меры точности реконструкции родословной является важной основой для сравнения различных конструкций записывающих устройств и для прогнозирования эффектов различных биологических факторов.
Все рецензенты прокомментировали значение показателей точности, задав вопрос о том, какой уровень точности потребуется для того, чтобы этот подход к генеалогии был полезным. Мы не можем дать единообразный ответ на этот вопрос, потому что требуемый уровень точности будет зависеть от задаваемых вопросов и биологических выводов, сделанных на основе клеточного происхождения. Исследователи должны будут оценить уровень точности происхождения, необходимый для ответа на заданный вопрос, в каждом конкретном случае.
Чтобы расширить и прояснить наши измерения точности происхождения, в исправленную рукопись мы включаем следующие новые результаты:
1) Помимо нашей глобальной меры точности, мы предоставляем оценки точности родословной на разных уровнях дерева родословной.Это дает представление о точности для больших (ранних) клонов по сравнению с более мелкими (более поздними) клонами.
2) Мы разработали новую меру точности реконструкции, которую мы можем применять на всех уровнях дерева родословных, чтобы дополнить нашу глобальную меру точности дерева. Этот показатель может показать, как информационное наполнение целей уменьшается во время разработки, когда цели становятся насыщенными.
3) Мы расширили оценки ложноположительных и ложноотрицательных результатов (показатель, который, по мнению обозревателей, полезен для понимания влияния точности) на все наши основные модели, и мы предоставляем оценки ложноположительных и ложноотрицательных результатов для различных размеров клонов.
Отдельных отзывов (просьба ответить по каждому пункту):
Рецензент № 1:
За последние несколько лет наблюдается бурный рост исследований по отслеживанию происхождения CRISPR. Однако оценить «точность» этих реконструкций родословной сложно, потому что правильная линия, как правило, неизвестна. В этом исследовании авторы в основном используют вычислительные подходы, наряду с некоторыми экспериментальными данными на эмбрионах дрозофилы, для оценки различных параметров, влияющих на точность реконструкции, таких как частота мутаций, скорость деления клеток, мутационное разнообразие и другие.
Некоторые мысли для авторов: что является достаточно точным с практической точки зрения?
Наша конечная цель – получить точное дерево родословной для всего эмбриона, но мы понимаем, что менее точные / разрешенные деревья также могут быть полезны, в зависимости от задаваемых биологических вопросов.
Мы поясняем это в отредактированной рукописи следующим образом:
1) Уже во введении мы заявляем: «Конечно, требуемая точность будет зависеть от предполагаемого использования линии передачи».
2) В Обсуждении мы добавили: «Требуемая точность линии передачи будет зависеть от приложения; например, точные деревья будут необходимы для обнаружения стереотипных делений и клеточных судеб, таких как те, которые обнаружены в предшественнике сенсорного органа Drosophila и линиях нейробластов ЦНС, но менее точные деревья могут быть достаточными для выявления предубеждений / тенденций, отражающих обязательства основных клонов ».
В эксперименте, где «настоящее» дерево неизвестно, как исследователи определяют соответствующие параметры?
В каком-то смысле это то, что мы делаем, поскольку дерево эмбриональных линий Drosophila неизвестно (на самом деле родословная не является стереотипной, она варьируется у каждого человека).Мы получаем как можно больше информации из экспериментальных данных и знаний о биологии мух и включаем эти параметры (скорость мутаций, мутационное разнообразие, скорость деления клеток) в наши модели для моделирования реконструкции этого неизвестного дерева.
В случаях, когда трудно определить скорость деления клеток, моделирование позволяет нам оценить ожидаемые уровни точности для ряда параметров. Как мы объясняем ниже (в ответ на пункт 5), мы обнаруживаем, что даже когда частота мутаций не может быть точно сопоставлена со скоростью деления клеток, реконструкция клонов может быть довольно точной при наличии достаточного количества мишеней.
Являются ли некоторые устройства записи CRISPR принципиально лучше, чем другие, и предлагает ли это исследование лучший подход?
Наше исследование предполагает, что конструкция регистратора важна, и что корректировка числа мишеней, частоты мутаций и количества выпадений будет иметь большое влияние на реконструкцию клонов с любым типом регистратора. Это сообщение четко прослеживается во всей рукописи.
Различные реализации записывающего устройства (GESTALT, MEMOIR, ScarTrace, LINNAEUS…), вероятно, будут отличаться по производительности, и это будет во многом зависеть от конкретных целевых чисел, частоты мутаций и т. Д.В этой рукописи мы специально тестируем работу двух опубликованных рекордеров (GESTALT и MEMOIR). Мы полагаем, что моделирование было бы полезным для тестирования любой конструкции записывающего устройства.
Основные комментарии:
1) Недавно было проведено систематическое наблюдение за van Overbeek et al., 2016, Leopold Parts в Sanger (Allen BioRxiv, 2018), в котором использовался большой набор данных для обеспечения инструмента прогнозирования распределения indel. Было бы очень интересно рассмотреть это для конкретных последовательностей-мишеней и обсудить «оптимизированные» спейсерные последовательности для регистраторов CRISPR, которые обладают более высоким мутационным разнообразием, чем другие.
Действительно, исследование Allen et al. (и два других недавно опубликованных исследования, Shen et al. и Chen et al.) предполагают, что на разнообразие и относительную частоту мутационных исходов влияют целевая и фланкирующая последовательности. Мы согласны с тем, что это будет важно учитывать при разработке последовательностей записывающих устройств. Эти прогнозы необходимо будет проверить экспериментально на соответствующих видах и в контексте геномного локуса, в котором находится регистратор CRISPR.
В отредактированном тексте мы подтверждаем это следующим образом: «Если, как и ожидалось, разнообразие мутаций и их относительная частота варьируются в зависимости от целевой последовательности и ее локальной среды (Overbeek et al., 2016; Vu et al., 2017; Allen et al., 2018; Shen et al., 2018; Chen et al., 2018) выборка различных целей для достижения этого оптимума была бы целесообразной ».
2) Обоснование предположения, что регистраторы CRISPR генерируют необратимые мутации-мишени, неясно.Например, может быть «обратная скорость», когда мутировавшая цель, которая создает одно основание, все еще может быть распознана Cas9 после деления клетки.
Насколько нам известно, не проводилось никаких систематических исследований, посвященных эффективности повторного нацеливания на мутированные сайты одной и той же гРНК. Однако мы ожидаем, что частота реверсий будет очень низкой, поскольку (1) после мутации большинство мишеней имеют множественные несовпадения с гРНК рядом с сайтом расщепления CRISPR (Allen et al., 2018), и ожидается, что эти сайты будут нацелены с очень низкой эффективностью по сравнению с немутантными мишенями (Hsu et al., 2013), (2) даже одиночные нуклеотидные несоответствия рядом с сайтом расщепления могут быть вредными для нацеливания CRISPR (см. Hsu et al. ., 2013 и наши результаты) и (3) даже когда мутировавшие мишени повторно расщепляются, подавляющее большинство не вернется к немутированной последовательности (см. Результаты мутаций в Allen et al., 2018). Основываясь на этих данных, мы оцениваем, что частота реверсии будет намного ниже, чем 1% от частоты прямой мутации.По этим причинам мы считаем, что скорость реверсии незначительна. «Необратимость» кажется разумным приближением при моделировании мутационного процесса и реконструкции клонов.
Мы добавили к Результатам предложение по этому поводу: «Мы ожидаем, что частота реверсий будет незначительной, поскольку даже однонуклеотидные изменения приводят к значительному снижению частоты мутаций».
3) Почему объединение соседей выбрано из соображений экономии (и из нескольких других возможных алгоритмов?). Этот вопрос следует подробнее рассмотреть.Могут ли авторы использовать свои идеи для улучшения этих алгоритмов реконструкции для решения проблем, связанных с регистраторами CRISPR?
Выбор NJ вместо Parsimony был практичным. Мы показали, что метод экономии в среднем будет немного точнее, чем в Нью-Джерси, однако он значительно медленнее. Теперь мы количественно определили разницу в скорости. В отредактированной рукописи мы показываем, что время, затрачиваемое методом экономии, экспоненциально увеличивается с глубиной клона, так что реконструкция клона всего из 180 клеток занимает ~ 52 часа (Рисунок 2 – приложение к рисунку 1B).С помощью NJ то же дерево можно восстановить менее чем за 1 секунду.
Хотя, возможно, стоит попытаться использовать экономию с реальным набором данных (вероятно, потребуется использовать двухэтапный эвристический подход и создать начальное дерево наилучшего предположения с использованием NJ с последующей оптимизацией с использованием критерия экономии и обмена ветвями), Для наших целей сравнения тысяч различных симуляций важно иметь хороший метод, а также иметь такой, который поддается вычислению.Большая часть нашего анализа касается сравнения эффектов различных дизайнов и различных параметров на точность реконструкции родословной, и выводы остаются в силе до тех пор, пока метод реконструкции дерева согласован.
В отредактированной рукописи мы обращаемся к этому вопросу следующим образом:
1) Мы приводим более подробное сравнение точности методов NJ и Parsimony (Рисунок 2 – приложение к рисунку 1A и B). Мы обнаружили, что Parsimony работает на 5-10% лучше, чем NJ для деревьев, содержащих до 180 ячеек.
2) Обосновываем выбор NJ в разделе «Материалы и методы».
4) Выбранное определение «точности» кажется проблематичным, потому что оно обычно не делает различий между ложноположительными и ложноотрицательными событиями реконструкции. Это кратко рассмотрено на рисунке 7, но его следует расширить.
Робинсона-Фулдса – это стандартная метрика, которая широко используется для сравнения деревьев в филогенетике. Мы приняли метрику, относящуюся к Робинсону Фулдсу, которая дает процент правильно решенных разделений в качестве глобальной меры точности дерева.Это подходит для сравнения общей точности реконструкции родословной, достигнутой в различных условиях. Это было разъяснено в рукописи.
Как было предложено, мы расширили использование ложноположительных и ложноотрицательных результатов – альтернативной меры точности дерева, предназначенной для более легкой интерпретации в контексте развития. Наше первоначальное использование ложноположительных и ложноотрицательных результатов рассматривало как клоны только одного размера. Теперь мы расширили подход FP / FN, чтобы охватить несколько клонов разного размера, и применили этот расширенный подход к моделированию рекордера CRISPR с 32 мишенями с оптимизированной частотой мутаций с отсевами и без них, к GESTALT и MEMOIR (раздел «Точность измерения» на разной глубине дерева »и рис. 9 в переработанной рукописи).
Наконец, чтобы обеспечить более полную оценку точности на разных глубинах дерева клонов, мы теперь записываем, как способность реконструировать клеточный клон меняется по дереву по мере того, как происходит процесс накопления мутаций (см. «Квартетный анализ»). Мы представляем этот анализ для моделирования рекордера CRISPR с 32 мишенями с оптимизированной частотой мутаций с отсевами и без них, для GESTALT и MEMOIR (рис. 8 в исправленной рукописи).
5) Авторы показывают, что установка правильной скорости мутации имеет значение.Как можно сопоставить частоту мутаций со скоростью деления клеток, если скорость или интервал клеточных делений неизвестен / сам по себе изменчив (например, в опухоли)? Об этом стоит поговорить.
Одним из важных результатов нашего моделирования было показать, что диапазон скоростей мутаций, которые производят аналогичную точную реконструкцию клонов, может быть довольно широким (см. Рис. 2C, D). Таким образом, даже когда частота мутаций не может быть точно согласована со скоростью деления клеток (например, когда они неизвестны или варьируются в пределах дерева), несоответствие между частотой мутаций и ожидаемой скоростью деления клеток будет иметь относительно ограниченное влияние на общую точность в довольно широких пределах. пределы.
Мы добавили к тексту следующее, чтобы прояснить его: «Диапазон скоростей мутаций, которые могут дать точную реконструкцию родословной, к счастью, оказывается довольно широким для данного размера дерева; Мутации от 0,05 до 0,25 на деление клетки могут дать достаточно высокий уровень точности для деревьев из ~ 65 000 клеток, если скорости деления относительно равны (рис. 2C, D). Такая гибкость будет полезна в случаях, когда скорость деления клеток плохо охарактеризована ».
Рецензент № 2:
Рукопись «Можно ли восстановить точную клеточную линию с помощью регистраторов CRISPR?» использует моделирование и некоторые экспериментальные данные для тщательного изучения параметров, определяющих точность отслеживания происхождения CRISPR.Они определяют и анализируют четыре ключевых параметра для двух типов регистраторов CRISPR: количество целей, частоту мутаций, состояния мутационных символов и отсевы. Эта работа важна и ценна для растущего сообщества, изучающего потенциал рекордеров CRISPR. Он обеспечивает путь для будущей оптимизации инструментов отслеживания происхождения этого типа. По большей части надписи и цифры четкие и информативные. У меня есть пять основных проблем, перечисленных ниже.
1) Провокационное название в форме вопроса несколько вводит в заблуждение – да? Рукопись не дает прямого ответа на этот вопрос.Я бы предложил более точное название, например «Конструктивные требования для более точных регистраторов CRISPR».
Мы думаем, что название рукописи («Можно ли восстановить точную клеточную линию с помощью регистраторов CRISPR?») Точно описывает вопрос, который мы рассматриваем в этой работе. Ответ на этот вопрос не является простым ответом «да / нет», а зависит от размера и сложности линии передачи, конструкции записывающего устройства и требуемого уровня точности.В нашей статье рассматриваются эти вопросы.
2) У меня есть две проблемы с термином «точность».
Первое беспокойство: авторы должны заранее четко указать свое определение термина «точность», которое включает как ложные срабатывания, так и ложноотрицательные результаты. Я бы сказал, что ложные срабатывания (неправильные точки ветвления) хуже, чем ложноотрицательные (т.е. отсутствующие точки ветвления). Разве в «точном» дереве не может быть недостающих точек ветвления, но нет неправильных точек ветвления? Авторы кратко описывают это различие, но только в самом конце раздела «Результаты», а затем вычисляют эти баллы «точности» только для подхода последовательного определения последовательности SOLID.Я был бы признателен за более подробное обсуждение этого определения и оценки в начале результатов, а также расчет FP / FN и отчет для всех симуляций.
См. Выше наш ответ по этому поводу рецензенту 1. Как объяснено в разделе «Материалы и методы» и показано на рисунке 9 – добавление к рисунку 1, как FP, так и FN получены из несоответствия между истинным деревом и реконструированным деревом. Ложноотрицательные ячейки не отсутствуют в дереве, а отсутствуют в реконструированной кладе / клоне, к которому они принадлежат (они неправильно размещены в другом месте дерева).
Вторая проблема: в целом, насколько точными нам нужны записывающие устройства CRISPR? Авторы установили высокую планку: полное и точное отслеживание происхождения 65 000 клеточного дерева. Достаточно ли 4% или 14% (окончательные «баллы», выставленные MEMOIR и GESTALT) для многих научных приложений? Это, безусловно, лучше, чем почти все существующие методы отслеживания происхождения, благодаря которым мы многое узнали о биологии. Я был бы признателен за обсуждение того, почему 100% полные и точные деревья являются такой важной целью, и чему мы все еще можем научиться у менее точных деревьев.
Мы согласны с тем, что для решения разных проблем требуются разные точности определения линий. Мы подробно обсуждаем этот вопрос и отвечаем на него в нашем ответе рецензенту 1 (выше).
3) Меня беспокоят три этапа подготовки библиотеки или фильтрации при секвенировании массива целевых сайтов Drosophila, которые четко не объяснены в тексте, что может уменьшить количество состояний символов, используемых во всех следующих имитациях:
a) Для вызова состояний символов – почему использовать только 9bp для фланкирования целевого сайта вместо простого выравнивания каждого чтения с неотредактированными сайтами? Я предполагаю, что это устранит некоторые состояния персонажа.
Действительно, мутации иногда выходят за пределы окна обнаружения в 9 п.н., поэтому важно спросить, было бы более полезным окно большего размера. Как указал рецензент 3, компромисс для увеличения окна обнаружения состоит в том, что больше ошибок последовательности будет захвачено и засчитано как ложные состояния символа.
Мы учли два момента, выбирая окно в 9 б.п. Во-первых, природа мутаций, индуцированных CRISPR, такова, что мутации сосредоточены в сайте расщепления.По нашим оценкам, мутации, выходящие за пределы этого окна обнаружения, имеют меньшую частоту, чем частота ошибок секвенирования (<0,4% мутаций пропускаются с окном 9 п.н.). Во-вторых, мутации, выходящие за пределы 9 п.н., представляют собой делеции (и несколько вставок), которые обычно вносят уникальные последовательности в окно обнаружения 9 п.н. (см. Allen et al. 2018). Мы ожидаем, что все эти изменения будут обнаруживаться как отдельные мутации.
Как предполагает рецензент, когда мы рассматриваем только 9 нуклеотидов, мы не можем отличить некоторые различные мутации друг от друга.Это может повысить уровень гомоплазии и снизить точность.
Наш анализ мутаций, зафиксированных при чтении 18 п.н. по сравнению с 9 п.н., показал, что мы неправильно группируем 4 пары состояний символов. Однако чтение 18 п.н. вносит дополнительные ошибки секвенирования: при чтении 18 п.н. ошибка секвенирования увеличится с 1,1% до 2,2%.
Чтобы количественно оценить общий эффект на точность определения линии, мы сравнили точность реконструкции линии с использованием 9 п.н. с тем, что мы получили бы с использованием 18 п.о. (оба кодируются как 60-символьные состояния).Средняя точность улучшается незначительно, с 72,19% до 72,24%.
Эти результаты включены в исправленную рукопись на рис. 4 – приложение к рисунку 1.
b) Если я понимаю фиг. 4A, авторы используют праймер, который находится непосредственно на результатах мутации PAM, которые нарушают любые основания внутри PAM, вероятно, не будут захвачены с помощью ПЦР и секвенирования. Могут ли авторы обсудить влияние этого?
Мишени были амплифицированы и секвенированы с использованием праймеров, расположенных на расстоянии> 40 нуклеотидов от каждого сайта-мишени.«Праймер», показанный на фиг. 4A, является неправильным и был удален. Приносим извинения за эту ошибку.
В разделе «Выделение ДНК, создание библиотек и секвенирование» Материалов и методов мы более четко объясняем использование праймеров для амплификации и секвенирования.
c) Зачем объединять 140 состояний редких символов в общее состояние (состояние 60)? Это, очевидно, приведет к ошибкам в дереве. Почему бы не относиться к ним так же, как к другим состояниям персонажей? Авторы пишут, что это было «для удобства», но я не уверен, почему это удобно, и это может быть потенциальным источником ложных срабатываний.
Я считаю, что в других документах показано большее количество потенциальных состояний персонажа, и я обеспокоен тем, что перечисленные выше шаги могут повлиять на моделирование.
Как мы упоминали в разделе «Реконструкция деревьев родословной с помощью объединения соседей (PAUP *)» в материалах и методах, максимальное количество различных состояний символа, которые мы могли использовать в программе анализа PAUP *, составляло 64.
Мы можем показать, что есть убывающая отдача от использования большего разнообразия состояний характера; например, сравните очень похожие результаты при рассмотрении состояний 16 и 32 символов на рисунке 2C.Мы проверили это дальше, сравнив наши результаты с 60 мутировавшими состояниями символов со случаем, когда мы сокращаем данные до 40 состояний символов (собирая вместе результаты с наименьшей частотой). Точность оптимальной реконструкции изменяется с 72% до 71% (Рисунок 4 – приложение к рисунку 1).
4) Авторы, по их собственному признанию, используют «самую пессимистичную оценку» частоты отсева. Вполне возможно, чтобы два разреза в пределах клеточного цикла приводили к двум изменениям вместо исключения.Рассматривали ли авторы явное использование своих собственных экспериментальных данных (даже с оговорками о подготовке библиотеки) или данных GESTALT для моделирования показателей отсева? Поскольку это предположение сильно влияет на точность, казалось бы, важно соблюдать осторожность при моделировании отсева.
Мы смоделировали GESTALT на основе экспериментальных данных. Когда мы моделируем наихудший сценарий для выбывших из школы, мы наблюдаем очень похожее количество аллелей на экспериментальные данные, предполагая, что реальность приближается к наихудшему случаю (см. Рисунок 6 – приложение к рисунку 1).
Для Drosophila у нас нет экспериментальных данных относительно отсева, но мы предоставляем результаты, охватывающие как лучшие, так и худшие сценарии.
5) Выбор алгоритма восстановления дерева имеет значение. В их собственном моделировании Parsimony оказывается более успешным, чем Neighbor Joining (рисунок 2 – приложение к рисунку 1). Однако в тексте об этом никогда не говорилось явно, и не обсуждается выбор алгоритма и его влияние на точность. Хотя штат Нью-Джерси был выбран по очевидным причинам (скорость), авторы должны подробно обсудить другие варианты и их влияние на точность дерева.
Мы обращаемся к этому моменту в нашем ответе рецензенту 1. Мы добавили новые результаты на рис. 2 – приложение к рисунку 1A и B, а также явное заявление в раздел «Материалы и методы».
Незначительные комментарии:
1) В подходе GESTALT с культивированием клеток также использовалась аналогичная стратегия набора нецелевых и 1 направляющей РНК, и на это следует ссылаться при обсуждении конструкции массива мух вместе с предостережениями, связанными с этим подходом (плохое редактирование на многих нецелевых объектах). ).
Мы добавили эту цитату в отредактированную рукопись: «Мы решили скорректировать частоту мутаций, изменив последовательность-мишень, чтобы ввести несовпадения в пару sgRNA: мишень (аналогично стратегии, используемой на культивируемых клетках McKenna et al., 2016) ».
Мы не считаем плохую скорость редактирования некоторых целевых вариантов проблемой. Цель этого массива состояла в том, чтобы измерить фактическую частоту мутаций на различных мишенях, что позволило нам выбрать мишени, которые соответствуют оптимальным скоростям для реконструкции клонов.Фактические регистраторы включают в себя массивы выбранной цели и не содержат субоптимальных целей.
2) В документе MEMOIR подробно рассматривается точность реконструкции дерева, включая сравнение с эталонными деревьями. Эти данные и обсуждения должны быть упомянуты в этом документе.
Мы благодарим рецензента за указание на это. Мы включили ссылки на Frieda et al., 2017 (MEMOIR), Schmidt et al., 2017 и Spanjaard et al., 2018 в исправленную рукопись (Введение, абзац пять).
Рецензент № 3:
В этой рукописи Сальвадор-Мартинес и Грилло и др. представить имитационное исследование недавно разработанных технологий отслеживания происхождения CRISPR. Авторы хорошо справляются с постановкой задачи, объясняя свой выбор различных параметров и допущений и добавляя экспериментальные данные на Drosophila, чтобы подкрепить этот выбор. Эта работа станет ценным дополнением к быстро развивающейся области, особенно в качестве реальной проверки степени инженерии организмов, которая, вероятно, потребуется для получения почти полных, точных деревьев с помощью этого класса методов.Наш первый важный комментарий касается тона статьи, в то время как наши дополнительные важные комментарии в первую очередь касаются альтернативных мер, которые должны быть оценены, которые обеспечивают больший баланс между строгой точностью и общим сохранением топологии дерева. Кроме того, перед публикацией необходимо устранить некоторые основные недостатки в дизайне экспериментов и анализе результатов редактирования у Drosophila.
1) Первый важный комментарий заключается в том, что мера точности, используемая в статье, очень консервативна, и в целом тон, который используется во многих частях статьи, (мы считаем) чрезмерно консервативным.Хотя строгая точность и полнота – это, конечно, цели, к которым стоит стремиться, они не являются предпосылками подобных экспериментов для достижения биологических открытий. Например, родственные отношения между клеточными типами могут быть точно выведены из общей топологии дерева, которое содержит неточности или неопределенность около его вершин.
Как обсуждалось в нашем ответе рецензенту 1, мы постарались внести ясность в отношении того факта, что требуемый уровень точности будет зависеть от приложения.
Одним из наиболее важных выводов нашего исследования является желательность проведения такого анализа мощности в качестве инструмента для оптимизации плана эксперимента и для оценки надежности конечного результата.
Для любой новой технологии исследования концепции – это всего лишь доказательство концепции, и всегда было ясно, что потребуется (и все еще требуется) значительный дополнительный инжиниринг, чтобы максимизировать ценность этих методов. Этот момент не умаляет ценности моделирования, представленного в этой статье.Более оптимистичный подход к тем же результатам заключается в том, что можно реконструировать большие деревья с достаточно высокой точностью (отлично!), Но для этого потребуется введение как минимум 50 целей (а в идеале несколько сотен целей), настройки скорости мутации. (хотя широкое плато, представленное на рисунке 2D, довольно обнадеживает), и тщательное рассмотрение переменных скоростей деления клеток. Эти выводы и другие анализы, представленные в документе, служат важным руководством для данной области (и проверкой реальности на фоне краткосрочного мышления), но статья часто принимает негативный тон, который, на наш взгляд, несовместим с самими результатами (например,грамм. тот факт, что определены условия, при которых достигается реконструкция с точностью 99%; подраздел «Оптимизация реконструкции клеточного клона для секвенирования in situ с 2-, 4- или 16-символьными состояниями», абзац шесть). Мы призываем авторов: (а) прояснить с самого начала статьи, насколько точность, как определено здесь, является весьма консервативным определением по сравнению с тем, что может потребоваться для получения биологической информации по деревьям, реконструированным с помощью ГЕСТАЛЬТА или связанные методы.
См. Наш ответ рецензенту 1.
(b) звучит более уравновешенно, с меньшим акцентом на то, что невозможно использовать системы, как указано в их пилотных реализациях, и больший акцент на пути вперед, то есть степени, в которой дальнейшее проектирование (больше целей , уменьшение межсайтовых делеций, настройка скорости мутаций и т. д.), чтобы получить максимальную отдачу от этих методов.
В рукописи оценивается точность текущих методов, не комментируя, что возможно, а что нет.Мы подробно обсуждаем способы улучшения регистраторов CRISPR и количественно оцениваем ожидаемые улучшения производительности. Мы пришли к выводу, что методы на основе CRISPR должны в конечном итоге позволить нам создавать точные деревья происхождения.
Последний абзац рукописи был изменен, чтобы дать четкий положительный ответ на вопрос, поставленный в заголовке: «Этот подход должен стимулировать разработку нового поколения регистраторов CRISPR, производительность которых будет явно оцениваться и адаптироваться для соответствия биологическим характеристикам. под рукой.Наше исследование предсказывает, что оптимизированные регистраторы в конечном итоге позволят реконструировать точные клеточные линии сложных многоклеточных организмов на уровне одной клетки ».
2) Моделирование технологий отслеживания происхождения на основе CRISPR в первом разделе (рис. 2) фокусируется на точности с помощью Робинсона-Фулдса, но должно включать характеристику с измерениями клональной реконструкции (анализ FP / FN). Эти измерения используются позже в разделе моделирования MEMOIR, и многие люди будут заинтересованы в использовании технологий отслеживания происхождения для этой цели.Упоминание или сравнение с другими показателями расстояния может быть более подходящим, см. Https://www.ncbi.nlm.nih.gov/pubmed/21383415 или обзор https://www.ncbi.nlm.nih.gov/pubmed/ 25378436
Как обсуждалось в ответах рецензентам 1 и 2, теперь мы расширили использование ложноположительных и ложноотрицательных результатов. Хотя существуют другие меры сходства деревьев, мы ожидаем, что их использование не изменит наших выводов относительно оптимального значения для данного параметра.
3) Вы показываете более высокую точность с максимальной экономией, но используете соединение соседей по всей статье.Важно убедительное обоснование этого выбора, поскольку это, по-видимому, сильное предубеждение против методов, которые вы оцениваете.
Мы рассмотрели это в нашем ответе судье 1 (пункт 3).
4) Робинсон-Фулдс – это метрика расстояния, и здесь вы нормализовали ее с точностью более [0–100%], которая нигде в статье не подробно описывается. Подробности в разделе «Материалы и методы» улучшат ясность статьи.
Теперь мы уточнили, что наша мера связана с метрикой расстояния Робинсона-Фулдса, но отличается от нее.Мы заявляем: «Точность реконструкции происхождения каждой симуляции определялась путем сравнения выведенного дерева с эталонным деревом с использованием меры, полученной из алгоритма Робинсона-Фулдса (Робинсон и Фулдс, 1981), который вычисляет процент расщеплений в эталонном дереве. дерево, которые точно восстанавливаются в выведенном дереве (рис. 2B). Если выведенное дерево идентично опорному дереву, точность Робинсона-Фулдса составляет 100% ».
В разделе «Материалы и методы» мы заявляем: «Точность реконструкции каждой клеточной линии была определена путем вычисления меры, связанной с расстоянием Робинсона-Фулдса (RF) между эталонным и предполагаемым деревьями.Мы подсчитываем процент разбиений (наборов ячеек, разделенных на две группы узлом в дереве) в ссылочном дереве, которые также встречаются в предполагаемом дереве ».
5) Подсчет 9меров – это немного либерально для определения результатов редактирования на основе целевой последовательности FAST. Учитывая известное расположение двухцепочечных разрывов и результаты восстановления, большинство результатов мутаций должно быть сосредоточено в разрезе или перекрывать его. При включении всех 9 проксимальных оснований захваченное разнообразие редактирования будет включать ошибки секвенирования и ошибки ПЦР.Это не должно повлиять на общий профиль результатов редактирования, но увеличит количество мутаций, и об этом следует упомянуть. Об этом свидетельствует повышенная частота мутаций в неармеченном столбце дополнительной таблицы 1 в дополнительном файле 1, поскольку некоторые из более активных нецелевых объектов имеют изменения в основаниях, которые, как считается, сильно препятствуют связыванию Cas9 (основания, которые очень близки к последовательности PAM. ).
Как мы отмечаем в нашем ответе рецензенту 2 (пункт 3a), существует компромисс при установке размера окна обнаружения между фиксацией мутационного разнообразия и исключением ошибок секвенирования.
Мы непосредственно количественно оценили частоту ошибок ПЦР / секвенирования у мух, не несущих Cas9 (показано как «нецелевое» в дополнительной таблице 1 в дополнительном файле 1). Для цели FAST этот показатель составляет 1%, по сравнению с целевым мутагенезом CRISPR 87% у эмбрионов и 92% у взрослых.
Мы изменили следующий текст в разделе «Результаты», чтобы сделать его более явным: «Как и ожидалось, мишень, которая имеет полную комплементарность с sgRNA Emx1.6 (мишень 16, названная мишенью« FAST »), показала самую высокую скорость мутаций; исправив ошибки секвенирования (~ 1% контрольных мишеней имеют различия из-за ошибок ПЦР или секвенирования, дополнительный файл 1), мы обнаружили, что 87% мишеней несли мутации в конце эмбриогенеза.”
6) Кроме того, расположение праймера (рис. 4) не позволяет найти делеции, которые простираются ниже по течению в последовательность PAM. Это исключает обнаружение мутаций, которые простираются на 3 фута вырезанной площадки, и снижает разнообразие редактирования, которое вы видите. Это серьезная ошибка, которая также может повлиять на результаты FAST, выходящие за рамки цели.
Как указано в нашем ответе рецензенту 2 (пункт 3b), на рисунке 4 была ошибка, которую мы сейчас исправили. Мишени были амплифицированы и секвенированы с использованием праймеров, расположенных на расстоянии> 40 нуклеотидов от каждого сайта-мишени, поэтому мы смогли найти все делеции, а не только те, которые находятся в окне 9 пар оснований.
7) Было показано, что частота очень больших делеций уменьшается с увеличением расстояния между двумя сайтами (https://www.ncbi.nlm.nih.gov/pubmed/24
3). стоило бы включить в моделирование отсева или, по крайней мере, упомянуть альтернативную модель с уменьшающейся эффективностью отсева на больших расстояниях.
Действительно, помимо скорости расщепления CRISPR, на частоту выпадения будет влиять взаимодействие нескольких параметров, таких как расстояние между участками разреза, скорость восстановления и т. Д.В настоящее время трудно предсказать, как эти параметры влияют на процент отсева, и, вероятно, они будут варьироваться в зависимости от вида и типа клеток. Данные, представленные в цитируемой статье, относятся к вариациям в скорости делеции на больших расстояниях (несколько т.п.н.) в клетках млекопитающих. Это не кажется особенно актуальным для моделируемых нами регистраторов CRISPR, которые намного меньше по размеру (расстояния между целями 100 п.о. для наших целевых массивов FAST и 26 п.н. для GESTALT).
Поскольку показатели отсева трудно предсказать, наш анализ 32 целей охватывал два условия: лучший сценарий без выпадения и наихудший сценарий с выпадением для каждого предсказанного двойного расщепления.Для GESTALT наихудший сценарий резюмирует имеющиеся экспериментальные данные.
8) Было бы хорошо охарактеризовать точность деревьев с разрушенным генотипом (https://www.ncbi.nlm.nih.gov/pubmed/29474671), которые стремятся уменьшить количество ложных точек ветвления (точки ветвления, представленные требование раздвоения дерева, а не мутации).
Это интересная идея и одна из нескольких, которые заслуживают рассмотрения в будущем для точной реконструкции данных о происхождении, но мы чувствуем, что это выходит за рамки данной рукописи.
9) В разделе моделирования SOLiD (для рисунка 7) неясно, какая последовательность будет использоваться для праймера и как часто этой связывающей последовательности будут препятствовать делеции в ее целевой последовательности. Некоторые подробности здесь были бы полезны.
В наших симуляциях SOLiD мы предположили, что праймер секвенирования никогда не будет потерян в результате мутации, поэтому наше моделирование представляет собой лучший сценарий. Чтобы минимизировать гомоплазию при захвате одного или двух прочтений SOLiD, мы обнаружили, что праймер должен быть оптимально размещен на 3 ’мишени, перекрывая PAM.
Этот эксперимент моделирует дополнительные ограничения, накладываемые существующей технологией для секвенирования на месте, и подчеркивает необходимость учета факторов, которые могут повлиять на количество и качество информации, исходящей из эксперимента.
Мы прояснили эксперимент SOLiD, добавив следующие предложения в раздел «Материалы и методы»: «Для моделирования секвенирования SOLiD мы использовали реальное распределение динуклеотидов, наблюдаемое в положениях 6-7 и 11-12 перед PAM.Мы преобразовали их в четыре различных цветовых состояния для каждого динуклеотида в соответствии с протоколом SOLiD (рисунок 7 – приложение к рисунку 1) ».
Незначительные комментарии:
Четвертое предложение реферата, вероятно, должно говорить о подходах (вы указываете как минимум два).
Изменено на «Здесь мы используем компьютерное моделирование для оценки эффективности этих подходов в различных условиях».
Abstract: не все терминальные ветви являются полностью дифференцированными клетками.
Изменено на «Деления, которые генерируют эти взрослые клетки, составляют генеалогическое дерево с оплодотворенной яйцеклеткой в его корне и каждой взрослой клеткой в качестве конечной ветви».
Введение, абзац второй: вероятно, есть лучшая фаза, чем «простые дела» (это были довольно трудные усилия, о которых вы упоминаете далее).
Изменено на «Получение клонов с высоким разрешением (на уровне одной клетки) – сложная задача, которая была решена только на животных с относительно небольшим количеством клеток, таких как нематода Caenorhabditis elegans : ее полное происхождение (~ 1000 клеток) было установлено кропотливое наблюдение за каждым делением клетки под микроскопом.”
Вступительный абзац восьмой немного резок. Конечно, моделирование будет информировать будущие синтетические регистрирующие системы, но допущения и упрощения моделирования могут помешать нахождению оптимального решения без более точной биологической проверки.
Изменено на «В конечном счете, это моделирование поможет нам установить набор критериев для оптимальной конструкции регистраторов клональных линий на основе CRISPR, а также понять ограничения этих методов при решении реальных биологических вопросов.”
Подраздел «Оптимизация реконструкции клеточного клона для секвенирования in situ с 2, 4 или 16 символьными состояниями», абзац девятый: не следует «удвоить количество считываний на цель» быть «удвоить длину считывания» или «удвоить количество циклов» ?
Изменено на «Очевидно, что увеличение количества мишеней улучшит производительность, но мы хотели знать, не лучше ли вместо этого читать удвоенное количество нуклеотидов на мишень, что представляет собой те же усилия по секвенированию.”
Действительно красивые рисунки на протяжении всего дополнения, но самое незначительное изменение, добавление «+ theme_classic ()» или аналогичное к команде R для рисунка 2 – приложение к рисунку 1.
Цифра изменена.
Дополнительные сведения необходимы в экспериментальном разделе о реакции ПЦР.
Мы добавили дополнительные сведения для решения этого вопроса.
https://doi.org/10.7554/eLife.40292.024Стал ли вирус Западного Нила забытой тропической болезнью?
Образец цитирования: Ronca SE, Ruff JC, Murray KO (2021) 20-летний исторический обзор вируса Западного Нила с момента его первоначального появления в Северной Америке: превратился ли вирус Западного Нила в забытую тропическую болезнь? PLoS Negl Trop Dis 15 (5): e0009190.https://doi.org/10.1371/journal.pntd.0009190
Редактор: Педро ФК Васконселос, Instituto Evandro Chagas, БРАЗИЛИЯ
Опубликован: 6 мая 2021 г.
Авторские права: et © 2021 Ronca al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Авторы были частично профинансированы NIH / NIAID (U19AI089992-06) (K.O.M.), Фондом Чао (K.O.M.) и Фондом Брокмана (https://brockmanfoundation.org) (K.O.M). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Интродукция и история вируса Западного Нила в Северной Америке
Вирус Западного Нила (ВЗН) был впервые обнаружен в Уганде в 1937 году [1] и более 60 лет циркулировал в цикле передачи энзоотических комаров по всей Африке, Ближний Восток, Россия и Европа, с преобладающим штаммом линии 2 [2].Инфекции обычно характеризовались как субклинические или вызывающие легкое лихорадочное заболевание [3]. В середине 1990-х годов появился новый штамм WNV (линия 1), который привел к высокой доле неврологических инфекций с эпизоотиями в Румынии, других частях Европы, России и Израиле [2,4].
В конце августа 1999 года врач-инфекционист (доктор Дебора Аснис) из района Квинс сообщил в Департамент здравоохранения и психической гигиены Нью-Йорка (NYCDOH) 2 случая энцефалита, что привело к расследованию [5] .Подобные случаи также были быстро выявлены в соседних больницах, и NYCDOH обратилось за помощью в Центры по контролю и профилактике заболеваний (CDC), чтобы помочь определить причину. На основе опросов пациентов и членов их семей, а также экологических проверок стало очевидно, что наиболее вероятным возбудителем болезни были комары. Первоначальный анализ сыворотки и спинномозговой жидкости (CSF) от подгруппы подозреваемых пациентов показал положительный результат на IgM-антитела против вируса энцефалита Сент-Луиса (SLEV) с помощью иммуноферментного анализа с захватом моноклональных антител (MAC-ELISA) в CDC. что привело к быстрому осуществлению мер по борьбе с комарами [6].
В то время как первоначально считалось, что массовая гибель американских ворон (семейство Corvidae) связана со вспышкой среди людей, патобиолог штата Нью-Йорк сообщил, что эти смерти были связаны с массовыми отравлениями. Примерно через 2 недели после начала расследования NYCDOH, содержащиеся в неволе экзотические птицы в зоопарке Бронкса начали умирать от энцефалита, который, как установил ветеринарный патолог зоопарка, доктор Трейси Макнамара [6], стал причиной смерти. Ткани головного мозга этих птиц были отправлены в лаборатории национальных ветеринарных служб Министерства сельского хозяйства США (NVSL), а вирусные изоляты затем отправлены в CDC для секвенирования [7].Секвенирование первоначально выявило штамм, наиболее вероятно связанный с WNV, первоначально называвшийся WNV-подобным, а затем подтвержденный как WNV линии 1, наиболее близкий к штамму Israel 1998 года [7]. Одновременно исследователи выделили вирус из мозга пациентов, умерших от энцефалита во время вспышки, и на основе секвенирования идентифицировали флавивирус, похожий на Кунджин / Западный Нил, что дополнительно подтвердило этиологию [8]. Затем было подтверждено, что смерть ворон была вызвана WNV [6]. К октябрю 15 лошадей заболели энцефалитом на Лонг-Айленде в Нью-Йорке и оказались положительными на ВЗН.Наконец, вирус был изолирован от комаров Culex pipiens и , что свидетельствует о предполагаемом переносчике вируса [9]. До 1999 года WNV никогда не обнаруживали в Западном полушарии.
Ретроспективный иммуноферментный анализ на IgM сыворотки и спинномозговой жидкости пациентов с энцефалитом или менингитом, у которых ранее проводился анализ на SLEV, был подтвержден положительным на инфекцию WNV, даже у тех, у кого ранее были отрицательные или сомнительные результаты SLEV [6,10]. В конечном итоге вспышка в Нью-Йорке (NYC) в 1999 г. привела к 62 подтвержденным случаям заболевания, включая 7 летальных исходов [5].Ближе к концу вспышки крупное кластерное серологическое обследование на уровне домохозяйств было проведено в наиболее пострадавшем районе Куинса [11]. Исследователи определили взвешенную (с поправкой на кластеры) серологическую распространенность инфекции WNV на уровне 2,6%. Основываясь на сообщенных признаках и симптомах участников исследования, исследователи экстраполировали, что примерно 80% инфицированных протекали бессимптомно, примерно у 20% развилось неосложненное лихорадочное заболевание, и только у 1 из 140 развился более тяжелый болезненный процесс в виде энцефалита или менингита, который позже был выдуман: « Нейроинвазивная болезнь Западного Нила »(WNND).По оценкам этого исследования, приблизительно 8 200 человек были инфицированы WNV в Нью-Йорке во время первоначальной вспышки [11].
До сих пор неизвестно, как вирус был занесен в Нью-Йорк. Правдоподобные теории включают занесение инфицированных комаров через морские перевозки или авиаперевозки, зараженных перелетных птиц из Европы, инфицированных завезенных птиц или домашних животных, преднамеренную интродукцию (биотерроризм) или виремическое лицо. Хотя люди считаются тупиковыми хозяевами, последняя теория могла бы быть возможной, если бы у инфицированного человека был серьезный иммунодефицит.
Географическое появление
После вспышки в 1999 году возникла неуверенность в способности вируса перезимовать, а затем распространиться в новые географические районы. Учитывая высокий процент смертей среди врановых, в Центре контроля заболеваний был установлен надзор за смертностью птиц в качестве средства раннего обнаружения вирусной активности и инструмента мониторинга географического распространения [12–14]. В конечном итоге, наблюдение за смертностью птиц было объединено с наблюдением за людьми, лошадьми и переносчиками в новый инструмент наблюдения под названием ArboNET, со связями карт в реальном времени, предоставленными Геологической службой США [15].Как и предполагалось, надзор за смертностью птиц стал эффективным инструментом раннего обнаружения по мере того, как он переместился в новые географические районы, причем обнаружение гибели птиц часто предшествовало обнаружению положительных случаев заражения комаров и людей [12,16–18].
В первый год после первоначальной вспышки в Нью-Йорке WNV оставался изолированным на северо-востоке, и 21 случай заражения людей был выявлен в Нью-Йорке, Нью-Джерси и Коннектикуте [19]. В 2001 г. WNV распространился географически вдоль восточного побережья: 66 случаев заболевания было выявлено в 10 штатах между Массачусетсом и Флоридой.При южном распространении вирус был обнаружен в популяциях Culex quinquefasciatus , что вызывает опасения по поводу дальнейшего эпидемического потенциала, как видно на SLEV, родственном флавивирусе [20–22]. Тем не менее, распространение и степень передачи эпидемии в 2002 году были беспрецедентными: 4 156 случаев заражения людей вирусом WNV в США, включая 284 смертельных случая [19], зарегистрированы на западе, в Техасе и Монтане, и на севере, в провинциях Квебек и Квебек. Онтарио, 414 зарегистрированных случаев заболевания в Канаде [23]. Эпизоотическая передача также была высокой: зарегистрировано более 15 000 случаев заражения лошадьми [24].Во время этой эпидемии быстро стало очевидно, что ЛЗН может передаваться от человека к человеку через донорство виремической крови, трансплантацию органов, трансплацентарное перемещение и грудное молоко [25]. Это побудило принять экстренные меры по разработке скрининга доноров крови для предотвращения заражения крови, который был осуществлен в 2003 году [26,27]. К 2003 году вирус был обнаружен в Culex tarsalis , и его беспрецедентное распространение продолжилось, охватив 45 штатов США, при этом было зарегистрировано 9862 случая и 264 смертельных случая [19].Важно отметить, что общее количество случаев заболевания в США в 2003 г. было несколько завышено по сравнению с предыдущими годами, поскольку CDC потребовали сообщать о случаях не нейроинвазивных заболеваний (например, лихорадки Западного Нила [WNF]) в органы здравоохранения [13 ]. Как и в США, WNV продолжил свое распространение в Канаде: на их веб-сайте здравоохранения сообщается о 5 провинциях с подтвержденными автохтонными инфекциями и 8 провинциях / территориях, в которых сообщается в общей сложности о 1481 заболевании людей. Серологические доказательства вируса были зарегистрированы у лошадей и птиц-хозяев в некоторых странах Латинской Америки и Карибского бассейна; однако случаи заражения людей были необъяснимо редкими [28].
К 2004 г. эпизоотия ВЗН распространилась по западному побережью США, а к 2012 г. все 48 континентальных штатов и округ Колумбия сообщили о местном заболевании человека [29]. В период с 2002 по 2007 год WNV находился на пиковом уровне эпидемии как в США, так и в Канаде (рис. 1), затем резко снизился в период с 2008 по 2011 год. В 2012 году неожиданная эпизоотия произошла с рекордным числом случаев в Техасе [30], Луизиане, Оклахома, Арканзас, Миссисипи и Алабама [19]. Большое количество случаев (> 2000 ежегодно) продолжало регистрироваться в период с 2013 по 2018 год, что свидетельствует о высоком потенциале эпизоотических вспышек в обозримом будущем.Факторы эпизоотической передачи, включая численность комаров, круговорот популяции птиц, а также климатические условия и изменение, требуют дальнейшего изучения, чтобы предсказать будущие вспышки.
Филогенетика
Сдвиг в циркулирующем штамме WNV наблюдался между первоначальной вспышкой в Нью-Йорке в 1999 г. и последующими вспышками с 2002 г. [2,20,31–34]. Штамм NY99 очень нейровирулентен у мышей [35]. При пассировании у хомяков этот штамм также вызывал хроническую почечную инфекцию, которая затем воспроизводилась у мышей с использованием штаммов после пассирования хомяка [36–39].Другой штамм, WN2002, с 2-мя нуклеотидными различиями, был идентифицирован во время вспышки 2002 г. и остается в обращении сегодня [21,34]. Как описано в 20-летнем анализе доступных генотипов WNV Hadfield и его коллегами, WN02 в сочетании с SW03 вытеснил штамм NY99 в США [20,21] (Рис. 2). Некоторые исследования предполагают, что виды комаров Culex более эффективно передают штаммы вируса WN02 по сравнению со штаммами NY99 [40,41], но не все исследования согласуются с этим [42–44].На моделях мышей было обнаружено, что этот штамм WN02 вызывает вялый паралич, подобный тому, что наблюдается при заболеваниях человека [35]. Постоянная оценка генотипа и фенотипа штаммов, выделенных из эндемичных регионов, необходима для обеспечения соответствия штаммов, используемых в моделях животных для оценки вакцин и терапевтических средств.
Инфекционное бремя WNV среди людей, США
За 20 лет существования WNV в США (с 1999 по 2019 гг.) В CDC ArboNET было зарегистрировано в общей сложности 51 702 случая WNV, в том числе 25 227 (48.8%) WNND и 2376 (4,6%) смертей [45]. Исходя из первоначальной оценки Мосташари 1 случая WNND на каждые 140 инфекций, эти 25 227 случаев WNND предполагают почти 3,5 миллиона случаев инфицирования в США на сегодняшний день. Однако дополнительные серологические исследования показывают, что эта экстраполяция, вероятно, занижена [46–48]. В исследовании, опубликованном в 2013 г. Петерсеном и его коллегами, оценивалась совокупная заболеваемость ЛЗН среди взрослых в США с 1999 по 2010 г., применяя методы Карсона и его коллег [47], и было установлено, что 3 миллиона взрослых, вероятно, были инфицированы ВЗН в течение этого периода времени [49]. .Поскольку примерно 40% всех случаев WNND произошли с момента исследования Петерсена, и их исследование не отражало инфекции у детей, наша команда обновила расчетное количество случаев во всех возрастных группах с 1999 по 2016 год, используя методы Петерсена, но включая детей, по оценке Мандалакас и его коллеги [48]. В этом исследовании мы оценили почти 7 миллионов случаев инфицирования WNV в континентальной части США. Это, вероятно, заниженная оценка, поскольку несколько исследований, оценивающих частоту тестирования, определили, что только примерно 40% случаев, соответствующих критериям WNND, проверяются при обращении в медицинское учреждение [50,51].
Понимание истинного бремени WNV имеет решающее значение. Определенные группы населения особенно уязвимы к инфекции, включая бездомных и других, на которых влияют социально-экономические детерминанты [52–56], а также тяжелые заболевания, в том числе пожилые люди и люди с сопутствующими заболеваниями [5]. Эпидемиологические исследования описывают тяжелые и хронические последствия инфекций WNV, особенно у лиц с WNND, когда до 40% пациентов не могут вернуться к своему исходному состоянию здоровья [57]. Понимание истинного бремени также предоставит ценные данные для оценки политики общественного здравоохранения по контролю и профилактике заболеваний, а также для определения потребности в инвестициях в клинические испытания для разработки вакцин и терапевтических средств.
Стоимость Западного Нила
В связи с высокой распространенностью и эндемичностью WNV в Северной Америке необходимо понимать, какое бремя несет система здравоохранения и какое влияние оказывает на экономику. Зограбиан и его коллеги определили краткосрочные затраты на ВЗН в Луизиане с июня 2002 г. по февраль 2003 г. в размере 10,9 млн долларов США, из которых 4,4 млн долларов США – медицинские расходы и 6,5 млн долларов США – немедицинские затраты, такие как потеря производительности [58]. В аналогичном исследовании Барбер и его коллеги [59] оценили затраты, связанные со вспышкой ВЗН в 2005 г. в округе Сакраменто, Калифорния.Эта вспышка среди 163 человек обошлась примерно в 2,98 миллиона долларов США, включая борьбу с переносчиками болезней (примерно 702 000 долларов США), медицинские расходы на лечение и снижение производительности. Они определили, что необходимо предотвратить только 15 случаев WNND, чтобы меры по борьбе с переносчиками считались рентабельными [59]. Исследование, проведенное в Квебеке, Канада, определило затраты региона с 2012 по 2013 год [60]. В этом исследовании 90 пациентов были оценены ретроспективно, и средние затраты составили 21 330 долларов США на пациента с энцефалитом, 8 124 долларов США на пациента с менингитом и 192 доллара США на пациента с WNF.В целом, по их оценкам, в 2012 году было израсходовано около 1,7 миллиона долларов США на 124 симптоматических случая и 430 000 долларов США на 31 симптоматический случай [60]. Пройдя немного дальше, Стейплс и его коллеги оценили как краткосрочные, так и долгосрочные затраты, связанные со вспышкой, охватившей 80 случаев в Колорадо в 2003 году. Важно выделить диапазон затрат для этих пациентов, поскольку затраты на неотложную помощь варьируются от примерно от 4000 до 325000 долларов США для случаев энцефалита, примерно от 5000 до 283000 долларов США для острого вялого паралича (ОВП), примерно от 1000 до 15000 долларов США для менингита и примерно от 500 до 24000 долларов США для WNF, поскольку это подтверждает крайний и уникальный сценарии, которые могут испытать пациенты.При оценке долгосрочных затрат диапазоны составляют примерно от 0 до 24 000 долларов США для случаев энцефалита (вероятно, из-за более высокого уровня смертности, чем другие), примерно от 600 до 440 000 долларов США для ОВП, примерно от 0 до 261 000 долларов США для менингита и примерно от 0 до 41 000 долларов США для WNF. Эти долгосрочные затраты включают в себя медицинские приемы и оборудование, лекарства, долгосрочное лечение и потерю производительности, но нам не хватает информации, чтобы понять, как параметры острой инфекции, такие как продолжительность пребывания в больнице, могут повлиять на долгосрочные затраты.Используя свои расчеты для этих пациентов, они оценили общую стоимость WNV с 1999 по 2012 год в США примерно в 778 миллионов долларов США с доверительным интервалом от 673 миллионов долларов США до 1,01 миллиарда долларов США [61]. Ежегодно это равняется 56 миллионам долларов США, потерянным WNV.
Исследование экономической эффективности вакцин, проведенное Зограбианом и его коллегами, показало, что универсальная программа вакцинации в США вряд ли приведет к экономии [62]. Однако это исследование было проведено в начале внедрения WNV и оценивало бремя случаев заболевания с 1999 по 2004 год.В то время было зарегистрировано примерно 16 000 случаев и 7 000 случаев WNND. Крупная вспышка 2012 г. надвигалась, но неожиданно. Авторы обсуждали, что риск заражения, вероятность симптоматического заболевания и стоимость вакцинации имеют решающее значение для оценки экономической эффективности. С тех пор как это исследование было опубликовано в 2006 году, эти факторы изменились. Кроме того, мы должны учитывать потребности в вакцинах в регионах с низкой плотностью населения, где борьба с комарами менее осуществима на больших территориях.
Трудно интерпретировать, повлияло ли это исследование напрямую на будущий прогресс разработки вакцин против WNV или на инвестиции в коммерциализацию имеющихся вакцин. Спустя более 10 лет, в 2017 году, Стейплс и его коллеги [61] оценили рентабельность целевой вакцинации и пришли к выводу, что вакцинация на основе возраста может быть наиболее экономически эффективным методом борьбы с WNV, но нам еще предстоит увидеть, чтобы вакцина достигла широкого распространения. рынок, чтобы оценить его в полной мере.
Разработка вакцины
Быстрое распространение WNV по Северной Америке в начале 2000-х годов и высокая стоимость заболеваемости вызвали большой интерес к разработке вакцины WNV как для людей, так и для других животных, особенно для лошадей.Инактивированная цельновирусная вакцина была быстро разработана и лицензирована для ветеринарного использования в 2003 г. [64]. В ближайшие годы были одобрены различные вакцины для лошадей, в том числе первая ДНК-вакцина, лицензированная Министерством сельского хозяйства США (USDA) в 2005 г. [64] (пресс-релиз: https://www.cdc.gov/media/ прессрел / r050718.htm).
Ряд кандидатных вакцин для человека все еще находится на доклинических стадиях разработки [63]. На сегодняшний день в ClinicalTrials зарегистрировано 9 клинических испытаний по оценке кандидатов вакцины против ВЗН для человека.gov, при этом были исследованы только 6 отдельных агентов (Таблица 1). До сих пор ни одно испытание на людях не продвинулось дальше фазы II.
Первым агентом, прошедшим клинические испытания фазы I, была живая аттенуированная химерная вакцина Западного Нила-Денге. Три исследования фазы I для этой вакцины-кандидата были зарегистрированы на сайте ClinicalTrials.govhttp: //clinicaltrials.gov/ в 2004, 2007 и 2014 годах. Эта химерная вакцина основана на вакцине-кандидате DENV-4 rDEN4delta 30 с prM и E гены белков заменены генами штамма WNVNY99 [65].Минимальные побочные эффекты наблюдались во время всех 3 испытаний фазы I. Исследования 2004 и 2007 годов продемонстрировали сероконверсию примерно у 75% вакцинированных участников в возрасте от 18 до 50 лет после одной дозы 10 3 или 10 4 бляшкообразующих единиц (БОЕ). Более высокая доза 10 5 БОЕ приводила к более низкому уровню сероконверсии (55%), но была увеличена до 89% после 6-месячной ревакцинации [65]. В исследовании 2014 года среди взрослых в возрасте от 50 до 65 лет у 19/20 (95%) субъектов произошла сероконверсия после одной дозы 10 4 БОЕ.Снова была введена 6-месячная ревакцинация, но она не привела к значительному увеличению титра антител и, следовательно, считалась ненужной [66].
В 2005 и 2006 годах в ходе двух испытаний фазы I началось изучение использования ДНК-вакцины, аналогичной той, которая была одобрена для использования на животных. Вакцины на основе нуклеиновых кислот кодируют специфические антигенные субъединицы патогена и привлекают клетки-хозяева для продуцирования этих антигенов. Это привлекательная технология, поскольку она может обеспечить длительный иммунитет без введения какой-либо части патогена, кроме антигена, тем самым обеспечивая специфический иммунный ответ без риска реактивации ослабленного живого вируса [67].Оба этих кандидата в вакцины кодируют одни и те же белки prM и E, используемые в химерном кандидате, описанном выше, хотя один включает модифицированную версию компонента промотора гена [67,68]. В обоих испытаниях сообщается о легких побочных эффектах, связанных с вакцинами, и благоприятных иммунных ответах. Более поздняя версия, которая включала модифицированный промотор гена, действительно вызывала более сильный клеточный и гуморальный иммунный ответ [68].
Кроме того, рекомбинантная субъединичная вакцина с адъювантом была зарегистрирована для клинических испытаний в 2008 году и запатентована Hawaii Biotech в 2018 году, хотя отчета о результатах клинических испытаний на людях не публиковалось.Другой, внесенный в список в 2015 году, исследовал инактивированную цельновирусную вакцину HydroVax-001, которая, как было обнаружено, в целом хорошо переносится при концентрации как 1 мкг, так и 4 мкг в последовательности из 2 доз. Как и ожидалось, с вакциной на основе инактивированного вируса ни у одного участника не было обнаружено виремии при тестировании через 4 дня после каждого введения, но сероконверсия через PRNT 50 при более низкой дозе не вызывалась, и только 31% сероконверсии произошел после более высокой дозы. Ответ, специфичный для ELISA, был немного лучше, достигая 41% сероконверсии после второй дозы 1 мкг и 75% после дозы 4 мкг [69].
Только 1 вакцина-кандидат вступила в фазу II клинических испытаний. Это агент ChimeriVax-WN002, еще один живой аттенуированный химерный вирус. Этот вирус также содержит гены белков prM и E из NY99, но построен на основе вируса желтой лихорадки 17D. Благоприятный профиль безопасности у взрослых был продемонстрирован в исследовании фазы I [70], а у пожилых людей – в 2 исследованиях фазы II [63,71]. Виремия была низкой и не ассоциировалась с нежелательными явлениями ни в одной возрастной группе. По крайней мере, 4-кратное увеличение титров антител наблюдалось у> 90% всех пролеченных участников в обоих исследованиях фазы II [63,71].
Эти предварительные испытания продемонстрировали, что все существующие вакцины-кандидаты в целом хорошо переносятся и обладают различной иммуногенностью, однако прогресс в направлении лицензирования для использования людьми со временем застопорился. Препятствия для продвижения этих испытаний могут включать финансовые проблемы и нормативные требования, которые может быть трудно оправдать из-за большого количества добровольцев и ресурсов, необходимых в случае спорадических вспышек WNV.
Терапия
Параллельно с задержкой утверждения вакцины также не существует специальных терапевтических средств для лечения инфекций WNV.В ряде серий случаев и тематических исследований описывается использование интерферона, рибавирина и кортикостероидов [72–77], но отчеты неубедительны, и никакие клинические испытания официально не оценивали эти варианты. Фактически, только 3 агента были зарегистрированы для клинических испытаний в США. В исследовании фазы I / II, зарегистрированном в 2003 году, изучали Omr-IgG-am, который представляет собой соединение внутривенного иммуноглобина (ВВИГ) с высоким титром против WNV, одобренное для использования в Израиле. В этом испытании была когорта с низкой и высокой дозой, и его контролировали с помощью активного плацебо IVIG (Polygam S / D) и пассивного плацебо в виде физиологического раствора.Не было отмечено значительных различий между группами лечения, активного плацебо или пассивного плацебо в отношении побочных эффектов или 90-дневного исхода. К сожалению, это испытание столкнулось с множеством проблем с набором участников и получением как исследуемого препарата, так и активного плацебо, поэтому оно было прекращено до достижения целевого набора, а группа лечения с более высокими дозами была прекращена [78].
Затем, в 2004 году, Sarepta Therapeutics зарегистрировала испытание по изучению фосфородиамидат-морфолиноолигомерного агента (PMO) AVI-4020.PMO представляют собой аналоги нуклеиновых кислот, которые в контексте WNV могут использоваться для блокирования трансляции вирусных белков, тем самым предотвращая репликацию вируса [79]. На момент написания этой статьи компания установила, что AVI-4020 присутствует в спинномозговой жидкости здоровых добровольцев в течение 18 часов после приема однократной дозы [80], но не было опубликовано никаких отчетов об испытании, посвященном изучению использования при WNND.
Последним агентом, зарегистрированным для клинических испытаний в США, является моноклональное антитело MGAWN1. Исследование фазы I с 2007 по 2009 год показало, что этот препарат в целом хорошо переносился 40 здоровыми участниками после однократного внутривенного введения в диапазоне от 0.От 3 мг / кг до 30 мг / кг. Было обнаружено, что у одного участника выработались антитела против MGAWN1 через 3 месяца после воздействия препарата. Авторы сообщают, что это могло повлиять на эффективность препарата для этого участника, но, поскольку разовая доза, по-видимому, обеспечивает достаточное покрытие на время острого заболевания, считается, что это не представляет большого риска для реакций чувствительности [81] . Учитывая очевидный успех исследования фазы I, в 2009 году началось исследование фазы II того же препарата.Однако, как и испытание OMR-IgG-am, оно было прекращено досрочно из-за низкой посещаемости. Все исследования сведены в Таблицу 2.
Одним из факторов, которые авторы исследования Omr-IgG-am называют препятствием для регистрации, был длительный период, необходимый для получения одобрения институционального наблюдательного совета (IRB) для каждого клинического центра. В настоящее время каждое клиническое учреждение, участвующее в исследовании, должно иметь независимое одобрение IRB, что может занять до 6 месяцев. Это исключает регистрацию любых острых случаев, которые присутствуют в больнице или клинике, которые еще не одобрены.Эти авторы утверждают, что наличие одного централизованного IRB облегчило бы своевременный набор участников из самых разных клинических центров [78]. Кроме того, спорадическое появление многих случаев WNND может затруднить достижение необходимой целевой регистрации.
Диагностика
Несколько исследований документально подтвердили недостаточную диагностику ЛЗН [27,50,82], при этом превалирующие и разнообразные препятствия для диагностики. У большинства инфицированных наблюдается легкое течение болезни с небольшими преходящими признаками и симптомами или их отсутствие (субклиническое заболевание) [83].Возможностей для диагностики этих случаев мало, поскольку их симптомы недостаточно серьезны для обращения за медицинской помощью. Эти пропущенные случаи не только усложняют расчет оценок заболеваемости и распространенности, но также вызывают озабоченность по поводу передачи через продукты крови, предоставленные бессимптомными виремическими донорами. В ответ на эту озабоченность в 2003 г. были инициированы требования по скринингу кровоснабжения США на вирусную РНК [26,27]. С тех пор эпиднадзор за предполагаемыми виремическими донорами крови со стороны общественного здравоохранения стал важным ресурсом для определения частоты бессимптомных и субклинических инфекций [46,84].
Даже клинически очевидные инфекции сталкиваются с проблемами диагностики. WNF – это слабо выраженный синдром, напоминающий грипп и другие вирусные синдромы [83,85]. Крупное исследование доноров крови в США показало, что только 38% субъектов, которые были положительными на ВЗН и имели симптомы, обращались за медицинской помощью, и только 5% из них имели формальный диагноз ВЗН [27]. Пациенты с нейроинвазивными проявлениями (WNND) чаще получают диагноз, чем пациенты с WNF, в основном потому, что серьезность признаков и симптомов требует медицинской помощи, а окончательный диагноз активно проводится.Клиническое тестирование на ВЗН проводится примерно в 40% случаев заражения ВЗН-совместимым менингитом и энцефалитом у взрослых [50,51] и примерно в 25% совместимых педиатрических случаев [50]. Пациенты с энцефалитом с большей вероятностью будут проверены на ВЗН, чем пациенты с менингитом, хотя, по оценкам, на менингит приходится от 30% до 50% случаев ЗНБ [50]. Недостаточная диагностика WNV дополнительно проиллюстрирована в многоцентровом анализе больничных тестов WNV среди пациентов с менингитом и энцефалитом.В этом исследовании 84% пациентов, у которых группа исследователей обнаружила, что они имеют положительный эффект на ВЗН IgM в спинномозговой жидкости, прошли тестирование на ВЗН в рамках диагностического обследования, но 25% пациентов, прошедших клиническое тестирование, были пропущены из-за неадекватного тестирования [82].
В настоящее время наиболее широко используемым методом диагностики ВЗН является обнаружение антител IgM к ВЗН в сыворотке или спинномозговой жидкости. У этого метода есть несколько важных недостатков. Во-первых, известно, что флавивирусы вырабатывают антитела, которые обладают перекрестной реактивностью с другими флавивирусами, и поэтому рекомендуется подтверждать все диагнозы, поставленные таким образом, путем отправки образцов сыворотки в остром периоде и в период выздоровления в референс-лабораторию для проведения тестов нейтрализации уменьшения образования бляшек (PRNT). ).Этот дополнительный этап должен выполняться в лаборатории с уровнем биобезопасности 3 (BSL-3) [86], что может быть неудобным и отнимать много времени. Альтернативно, для использования PRNT доступны псевдотипные вирусы, что дает возможность обойти необходимость сдерживания BSL-3 [87]. Во-вторых, антитела IgM появляются в сыворотке крови между 3 и 8 днями после появления симптомов, поэтому возможно, что раннее тестирование может дать ложноотрицательный результат. Антитела IgM к WNV могут сохраняться в сыворотке крови в течение многих лет после острой инфекции [88,89], при этом в одном исследовании сообщалось об обнаруживаемых уровнях IgM в течение 8 лет после инфицирования примерно у 20% участников [90].Кроме того, доступные в настоящее время тесты для обнаружения WNV IgM не обладают высоким уровнем чувствительности, только 54% для ELISA и 45% для иммунофлуоресцентного анализа (IFA), а показатели чувствительности и специфичности могут варьироваться в зависимости от вирусных клонов [91]. Это значительно снижает диагностическую ценность этого теста в острых условиях, поскольку нельзя быть уверенным в том, что отсутствие IgM исключает диагноз ЛЗН или его присутствие указывает на острую инфекцию.
Ограничения тестирования были важным фактором в обеспечении адекватного скрининга донорской крови.Тестирование нуклеиновой кислоты (NAT) донорской плазмы используется для скрининга донаций крови в США с 2003 года. Мини-бассейны, состоящие из нескольких донорских образцов, обычно тестируются в первую очередь (MP-NAT), а реактивные пулы дополнительно исследуются с помощью индивидуального тестирования донации (ID- NAT) каждого образца, включенного в пул. Поскольку ID-NAT более чувствителен, чем MP-NAT, существуют протоколы, позволяющие лабораториям переключаться на ID-NAT в условиях высокой активности WNV. За годы, прошедшие после внедрения этих методов скрининга, идентификация ряда связанных с трансфузией передач ЛЗН вызвала интерес к распределению вирионов в различных компонентах крови.В исследовании 2007 года, проведенном учеными Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), использовалась полимеразная цепная реакция с обратной транскрипцией (ОТ-ПЦР) для оценки вирусной нагрузки в компоненте красных кровяных телец (RBC) по сравнению с компонентом плазмы, и было обнаружено, что вирусная нагрузка составляет один порядок по величине выше в компоненте эритроцитов [92]. Другие продолжили это исследование и подтвердили, что РНК ВЗН присутствует в компоненте эритроцитов до 3 месяцев [93]. Интересные теории, возникшие в результате этого исследования, включают в себя то, что прикрепление вирусов к эритроцитам происходит примерно во время сероконверсии [86] и что группа крови может влиять на способность вирионов прикрепляться к белкам на клеточной мембране [93].Включение ПЦР цельной крови с тестированием на IgM может иметь решающее значение для своевременного и экономичного выявления случаев заболевания в будущем.
ОТ-ПЦР использовалась для идентификации РНК ВЗН в других типах образцов, включая спинномозговую жидкость, мочу и слюну [82,94–97]. Тестирование на вирусную РНК в спинномозговой жидкости, по-видимому, имеет ограниченную ценность, поскольку в некоторых отчетах утверждается, что его часто невозможно обнаружить во время появления симптомов [82]. Это может помочь объяснить, почему только 16,6% образцов от пациентов, которым был поставлен диагноз ЛЗН с помощью других методов, были положительными в спинномозговой жидкости после ОТ-ПЦР [97].На сегодняшний день только одно исследование изучило присутствие РНК ВЗН в слюне. Он был обнаружен только у 1 из 10 участников и сохранялся примерно до 9 дней после начала заболевания [95]. В нескольких исследованиях изучалась полезность тестирования РНК ВЗН в моче [94,95,97], и хотя он, по-видимому, работает лучше, чем слюна и спинномозговая жидкость, наилучшая зарегистрированная оценка чувствительности составила 58,3% [97]. Что касается тестирования цельной крови, результаты этих исследований согласуются с ранее процитированной работой. Было обнаружено, что цельная кровь имеет чувствительность 86.8% [97], а модель распределения Вейбулла, основанная на банках образцов из когорты Западного Нила Хьюстона, показала, что большинство серийных образцов цельной крови были положительными на РНК WNV примерно до 3 месяцев после появления симптомов (50% отрицательных на 79 день и 95% отрицательных на 119 день) [95].
Заболевание, которым не уделяют должного внимания: грантовое финансирование и публикации с течением времени
В течение последних двух десятилетий ВЗН обсуждается как развивающееся инфекционное заболевание, но мы и другие находим доказательства, подтверждающие его обозначение как забытое заболевание.Хотя исследования продолжаются, стандарты ухода по-прежнему ограничиваются только поддерживающими мерами без проверенных терапевтических целей или профилактических вакцин. Когда грантовое финансирование, направленное на эти усилия со стороны Национальных институтов здравоохранения (NIH), оценивалось с помощью инструмента RePORTER, около 67 миллионов долларов США было направлено на исследования, связанные с WNV с 2000 по 2019 год (примерно 3,4 миллиона долларов США в год), хотя некоторые из этого было посвящено общему исследованию флавивирусов. Количество заявок на гранты, финансируемых NIH в год с того времени, также может быть записано (рис. 3), при этом большинство этих грантов составляют R01, за ними следуют небольшие гранты, такие как R03 и R21, с наименьшим объемом финансирования, обнаруженным в развитии карьеры. награды.Для сравнения, за тот же период NIH присудил более 922 миллионов долларов США за исследования вируса Зика, причем более 99% наград было получено в период с 2016 по 2019 год (примерно 230 миллионов долларов США в год).
Поиск в PubMed исследования «Вирус Западного Нила» в заголовках проиндексированных статей дает 3978 статей (рис. 4). Эти статьи охватывают период с 1946 года по начало ноября 2019 года и посвящены статьям, в которых подробно исследуются аспекты WNV, будь то экология, эпидемиология, генетика, последствия, терапевтические разработки или другие важные аспекты процесса болезни.
WNV не только широко исследовался сам по себе, но также использовался в качестве инструмента или руководства для определения механизмов и исходов связанных флавивирусных инфекций. Фактически, если этот поиск PubMed будет расширен за счет включения «вируса Западного Нила» как в заголовки, так и в абстрактные поля, по состоянию на начало ноября 2019 года можно будет найти 6 229 результатов. Многие из этих дополнительных примерно 2000 статей ссылаются на пути или результаты WNV в качестве ориентира. принцип исследования, которое стремились завершить их команды.Интересно отметить, что при сравнении публикаций WNV с другим флавивирусом – вирусом Зика – за период с 2016 по 2019 год в заголовке примерно 3600 статей упоминается вирус Зика. меньше, чем у других, недавно возникших и родственных инфекций.
.